Химическая природа и влияние на обмен гормонов щитовидной железы
Нормальная функция щитовидной железы направлена на секрецию L-тироксина (Т4) и 3,5,3′-трийод-L-тиронина (Т3) — йодированных аминокислот, которые представляют собой активные тиреоидные гормоны и влияют на разнообразные метаболические процессы (рис. 324-1). Заболевания щитовидной железы проявляются качественными или количественными изменениями секреции гормонов, увеличением размеров органа (зоб) или тем и другим вместе. Недостаточность секреции гормонов приводит к развитию синдрома гипотиреоза, или микседемы, главной особенностью которого служит снижение калорических затрат (гипометаболизм). Напротив, чрезмерная секреция активных гормонов вызывает появление гиперметаболизма и других признаков синдрома, называемого гипертиреозом, или тиреотоксикозом. Увеличение массы щитовидной железы (составляющей у взрослого человека в норме 15-25 г) может быть диффузным или очаговым. Диффузное увеличение необязательно должно быть полностью симметричным. Обычно правая доля железы увеличивается больше, чем левая. Такое увеличение может сопровождаться повышенной, нормальной или сниженной секрецией гормонов, что определяется причиной заболевания. Истинно очаговое увеличение отражает, как правило, наличие новообразований, будь то добро- или злокачественных. Первые иногда обусловливают гиперсекрецию тиреоидных гормонов и гипертиреоз, тогда как при злокачественных опухолях это наблюдается очень редко. Зоб любого типа может привести к сдавлению соседних структур шеи и средостения.
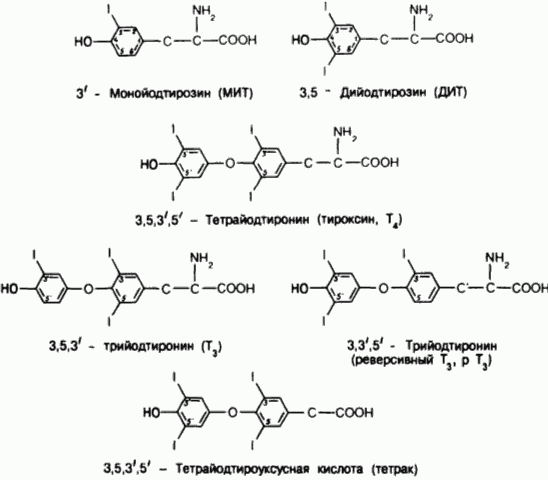 Структурные формулы тироксина, его предшественников и некоторых метаболитов.
Структурные формулы тироксина, его предшественников и некоторых метаболитов.
Гормоны щитовидной железы ( T3, T4, кальцитонин): Эмбриология, анатомия и гистология
Щитовидная железа человека развивается в эмбриогенезе из выпячивания глоточного эпителия и клеток латеральных глоточных карманов. Постепенно опускаясь по средней линии, зародышевая щитовидная железа формирует щитоязычный проток, который тянется от слепого отверстия у основания языка до перешейка железы. По ходу этого тракта могут сохраняться остатки ткани в виде «язычной щитовидной железы», щитоязычных кист и узлов или структуры, прилегающей к перешейку щитовидной железы и называемой пирамидальной долей. Последняя обычно видна только в случае увеличения остатка железы. У некоторых людей единственной функционирующей тиреоидной тканью может быть «язычная щитовидная железа», секреция которой бывает либо достаточной, либо недостаточной для сохранения нормального метаболического (эутиреоидного) статуса.
Аплазия щитовидной железы и функциональная недостаточность эктопической тиреоидной ткани служат причиной спорадического неонатального гипотиреоза, имеющего важное значение для здравоохранения вследствие частоты встречаемости (1 случай на 4000-5000 новорожденных) и способности отвечать на своевременно начатое лечение.
Щитовидная железа плода приобретает способность концентрировать и органифицировать йод примерно к 10-й неделе беременности. Вскоре после этого в крови уже удается определять как Т4, так и тиреотропный гормон (тиреотропин, ТТГ), концентрации которых на протяжении II триместра беременности непрерывно возрастают. Увеличение уровня Т4 в сыворотке крови плода обусловлено как усилением секреторной активности щитовидной железы и появлением в плазме тироксинсвязывающего глобулина (ТСГ), так и повышением уровня ТТГ, отражающим созревание гипоталамуса плода и секрекцию им тиреотропинрилизинггормона (ТРГ). Материнский ТРГ легко проникает через плаценту и, по-видимому, играет роль в развитии гипофизарно-тиреоидной системы плода.
Напротив, материнский ТТГ через плаценту не проникает. Т3 появляется в крови плода позднее, но также в течение II триместра, и его концентрация в крови и амниотической жидкости остается низкой вплоть до начала послеродового периода. В отличие от этого концентрация его аналога (реверсивного Т3, рТ3) в крови плода и амниотической жидкости превышает таковую в крови матери. Эти различия отражают качественные особенности метаболизма ‘T4 у плода, которые рассматриваются ниже. Низкая концентрация Т3 в крови плода и амниотическое жидкости на фоне его высокой концентрации у матери свидетельствует о минимальном переносе Т3 от матери к плоду, что характерно и для Т4. Следовательно, основным тиреоидным гормоном, получаемым плодом, является Т4, продуцируемый собственной щитовидной железой плода. Таким образом, за исключением возможного влияния материнского ТРГ, гипофизарно-тиреоидная ось плода представляет собой функциональную систему, независимую от таковой у матери.
Нормальная щитовидная железа у взрослого человека состоит из соединенных перешейком двух долей и располагается кпереди и книзу от хрящей гортани. Фиброзные перегородки делят железу на псевдодольки, которые в свою очередь состоят из везикул, называемых фолликулами, или ацинусами, окруженных сетью капилляров. В норме стенки фолликула выстланы эпителиальными фолликулярными клетками кубической формы. Просвет фолликула заполнен белковым материалом, получившим название коллоид, который содержит специфический для щитовидной железы белок — тиреоглобулин, ответственный за синтез и накопление Т4 и Т3. В щитовидной железе присутствует и другая популяция клеток — С-клетки. Они служат источником кальцитонина, а их злокачественное перерождение приводит к медуллярному раку щитовидной железы.
Динамика тиреоидных гормонов: нормальная физиология
Понятие «динамика тиреоидных гормонов» означает комплекс процессов синтеза гормонов в щитовидной железе, их транспорта в крови, действия и метаболизма в периферических тканях, а также комплекс регуляторных механизмов, определяющих нормальное обеспечение тканей тиреоидными гормонами.
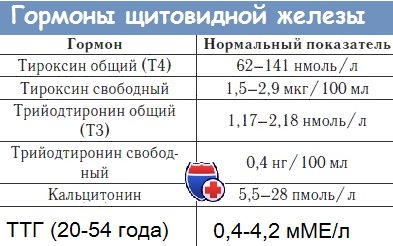
Гормоны щитовидной железы ( T3, T4, кальцитонин): Синтез и секреция гормонов.
Синтез тиреоидных гормонов зависит от поступления в щитовидную железу достаточного количества йода — составной части активных гормонов (Т4 и Т3), интактности путей метаболизма йода в железе и одновременного синтеза
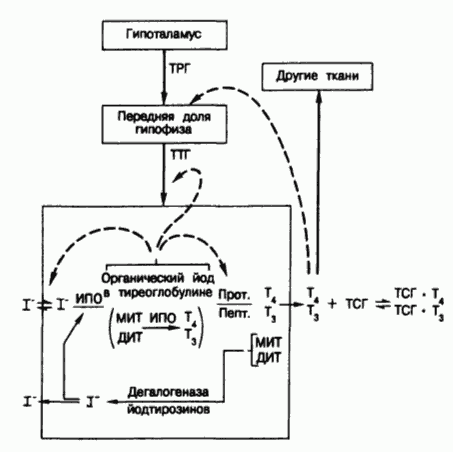 Схема путей синтеза и секреции тиреоидных гормонов и механизмов супра- и интратиреоидной регуляции функции щитовидной железы.
Схема путей синтеза и секреции тиреоидных гормонов и механизмов супра- и интратиреоидной регуляции функции щитовидной железы.
Тонкими стрелками показаны пути метаболизма йода; жирными стрелками — стимулирующие влияния; пунктиром — ингибирующие влияния.
Обозначения : ТРГ — тиреотропин-рилизинг гормон, ТТГ — тиреотропный гормон, И ПО — йодид-пероксидаза, Прот — тиреоидная протеаза, Пепт — тиреоидная пептидаза, МИТ — монойодтирозин, ДИТ — динодтирозин, T4 — тироксин, Т3 — 3, 5, 3′ — трийодтиронин.
Секреция достаточного количества гормонов требует в свою очередь, как нормальной скорости их синтеза, так и интеграции с протекающими в железе процессами гидролиза тиреогло-булина, в результате которых активные гормоны высвобождаются. Йод проникает в щитовидную железу из крови в форме неорганического или органического йодида. Существует два источника его поступления: первый — при дейодировании тиреоидных гормонов или насыщенных йодом агентов, попавших в организм человека; и второй — с пищей, водой или лекарственными препаратами. Раньше для населения континентальной части США считалось нормой потребление с пищей примерно 200 мкг йода; этого было достаточно для поддержания концентрации йодида в плазме па уровне приблизительно 0,5 мкг/дл (5 мкг/л).
Однако из-за присутствия йода в некоторых пищевых продуктах и широкого распространения йодсодержащих лекарственных средств, витаминных препаратов и антисептиков среднее потребление йода возросло до 1000 мкг в сутки, что привело к соответствующему повышению концентрации йодида в плазме крови.
Йодид извлекается из плазмы щитовидной железой, почками, а также слюнными железами и в желудочно-кишечном тракте, но, поскольку йодид, вы-деляющийся в просвет кишечника, подвергается реабсорбции, чистый его клиренс осуществляется только щитовидной железой и почками. В сущности, щитовидная железа и почки конкурируют друг с другом за йодид плазмы. Почечный клиренс зависит в основном от скорости клубочковой фильтрации, и на него не влияют гуморальные факторы или концентрация йодида в плазме. Поэтому почки в норме являются пассивными участниками этой конкуренции.
Отсюда следует, что соотношение между скоростью поступления йодида в щитовидную железу и скоростью его экскреции с мочой определяется активностью именно щитовидной железы, а не почек.
Процессы синтеза и секреции активных тиреоидных гормонов можно разделить на четыре последовательных этапа. Первый включает активный транспорт йодида из плазмы в клетку щитовидной железы и в просвет фолликула. Скорость этого процесса превышает скорость пассивной днффузии йода из железы. В результате щитовидная железа оказывается способной удерживать градиент концентрации для йодида (отношение концентраций щитовидная железа/плазма) на весьма высоком уровне (до 500 и более в определенных условиях). Энергия для транспорта йодида черпается из фосфатных связей и поэтому зависит от окислительного фосфорилирования в железе. Второй этап биосинтеза гормонов включает окисление йодида в более ре-акционноспособную форму, способную йодировать тирозиновые остатки в молекуле тиреоглобулина — гликопротеида с мол. массой около 650 000, который синтезируется клетками фолликулов.
Окисление йодида осуществляется йодид-пероксидазой, использующей перекись водорода, которая образуется но ходу окислительного обмена в железе. Йодирование органических структур происходит на границе между клеткой и коллоидом, где этому процессу подвергается в основном свежесинтезированный тиреоглобулин, поступающий путем экзоцитоза в просвет фолликула. В результате в составе пептида образуются неактивные предшественники гормонов — монойодтирозин (МИТ) и дийодтирозин (ДИТ). Затем эти йодтирозины вступают в реакцию окислительной конденсации опять-таки с помощью пероксидазы. Данная реакция протекает внутри молекулы тиреоглобулина и приводит к образованию различных йодтиронинов, включая Т4 и Т3 Хотя в крови и присутствуют небольшие количества тиреоглобулина, большая его часть некоторое время хранится в железе, играя роль запасной формы тиреоидных гормонов, или «прогормона». Высвобождение активных гормонов в кровь происходит путем пиноцитоза фолликулярного коллоида на апикальном краю клетки с образованием коллоидных капелек. Для этого процесса необходимо функционирование микротрубочек.
Коллоидные капельки сливаются с тиреоидными лизосомами, образуя «фаголизосомы», в которых тиреоглобулин гидролизуется протеазами и пептидазами. Конечный этап заключается в выделении свободных йодтиронинов — Т4 и Т3 — в кровь. Единственным источником эндогенного Т4 служит щитовидная железа. В отличие от этого только около 20% образующегося в норме Т3 поступает из щитовидной железы; остальная его часть образуется во внетиреоидных тканях путем ферментативного отщепления 5′-йода от наружного кольца молекулы Т . Неактивные йодтиро-зины, высвобождающиеся при гидролизе тиреоглобулина, отдают свой йод под действием внутритиреоидного фермента — дегалогеназы йодтирозинов. В норме высвобождающийся таким образом йод в основном реутилизируется в синтезе гормонов, но небольшая его доля все же теряется, поступая в кровоток («утечка йода»). В патоло-гических условиях эта доля может возрастать.
Щитовидная железа способна концентрировать и другие одновалентные анионы, такие как пертехнетат, который имеется в виде радиоактивного изотопа — натрий [99mТc] пертехнетат. В отличие от йодида пертехнетат очень мало связывается орга-ническими соединениями. Поэтому он присутствует в щитовидной железе только короткое время. Это свойство наряду с его коротким физическим периодом полураспада делает пертехнетат ценным радионуклидом для получения изображения щитовидной железы с помощью методов сцинтилляционного сканирования.
Перечисленные выше реакции служат объектом торможения различными химическими соединениями. Их обычно называют зобогенными веществами, поскольку в силу своей способности ингибировать синтез гормонов и косвенно стимулировать секрецию ТТГ они вызывают образование зоба. Некоторые неорганические анионы, в том числе перхлорат и тиоцианат, ингибируют механизм транспорта йодида и тем самым уменьшают доступность субстрата для образования гормонов. Однако развивающиеся в результате этого зоб и гипотиреоз можно предотвратить или ликвидировать достаточно большими дозами йодида, которые обеспечивают поступление нужных его количеств в железу за счет простой диффузии. Широко используемые антитиреоидные средства, такие как производные тиомочевины и меркаптоимидазола, оказывают на биосинтез гормонов более сложное воздействие.
Эти вещества, равно как и некоторые производные анилина, ингибируют первоначальное окисление (органическое связывание) йодида, снижая долю образующегося ДИТ относительно МИТ и блокируя конденсацию йодтирозинов в гормонально-активные йодгиронины. Последняя реакция наиболее чувствительна. Таким образом, синтез гормонально-активных йодтиронинов может быть резко заторможен в условиях лишь небольшого снижения общего захвата йода щито-видной железой. В отличие от эффекта одновалентных анионов зобогенное действие ингибиторов органического связывания йода не преодолевается большими его количествами. Действительно, некоторые слабые зобогенные вещества, такие как сульфона-миды и антипирин, при введении вместе с йодидом становятся почему-то даже более активными. Острое введение больших доз самого йода тоже может приводить к блокаде органического связывания и реакции конденсации. В норме это действие (эффект Вольффа — Чайкоффа) транзиторно, но у некоторых здоровых лиц, длительно получаю-щих йод, имеет место постоянное торможение синтеза гормонов, сопровождающееся развитием зоба с гипотиреозом (йодная микседема) или без него.
Большинство больных с болезнью Грейвса, особенно перенесшие радиойодтерапию или хирургическую операцию, а также больные с болезнью Хашимото чрезвычайно чувствительны к блокирующему действию йодида, и при хроническом приеме йодидов у них развивается гипотиреоз. Точно так же высокую чувствительность обнаруживает и щитовидная железа плода, и поэтому во избежание зобного гипотиреоза у плода беременные женщины не должны получать больших доз йодида. Йодид в больших дозах может ингибировать и протеолиз тиреоглобулина, т. е. высвобождение гормонов. Этот эффект легче всего проявляется в условиях гиперфункции щитовидной железы, и именно он определяет быстрое терапевтическое действие йодидов у большинства больных гипертиреозом. Литий, вводимый ряду больных с депрессивными состояниями в виде карбонатной со-ли. оказывает несколько эффектов на внутритиреоидный обмен йода, один из которых заключается в торможении секреции гормонов. Большие дозы дексаметазона также ин-гибируют секрецию гормонов и в сочетании с йодидом могут быстро уменьшать выра-женность тиреотоксикоза.
Гормоны щитовидной железы ( T3, T4, кальцитонин): Транспорт и метаболизм гормонов
Транспорт гормонов. В крови Т4 и Т3 почти полностью связаны с белками плазмы. В порядке уменьшения интенсивности связывания Т4 эти белки располагаются следующим образом: альфа-глобулин, называемый тироксин- или тиронин-связывающмм глобулином (ТСГ), Т4-связывающий преальбумин (ТСПА) и альбумин. Из-за своего вы-сокого сродства к Т4 СТГ в норме является главной детерминантой общей связывающей активности плазмы. Взаимодействие между Т4 и его связывающими белками формирует обратимое равновесное состояние, при котором большая часть гормона оказывается связанной, а очень малая его доля (в норме около 0,03%) — свободной. Т3 связывается с ТСПА в незначительной степени, а с ТСГ- менее прочно, чем Т4. Вследствие этого доля свободного Т3 в норме (примерно 0,3%) в 8-10 раз превышает таковую свободного Т4.
Ткани используют только свободный, или несвязанный, гормон. Поэтому метаболический статус теснее коррелирует с концентрацией именно свободного гормона, чем с общей его концентрацией в плазме, а гомеостатическая регуляция тиреоидной функции тоже направлена на поддержание нормальной концен-трации свободного, а не общего гормона. Кроме того, относительно слабое связывание Т3 обусловливает незначительность его вклада в общую концентрацию белково-связанного гормонального йода в крови и, возможно, более быстрое начало и окончание его действия. Нарушения взаимодействия между тиреоидными гормонами и бел-ками плазмы бывают двух общих типов.
Таблица. Классификация различных нарушений взаимодействия тиреоидных гормонов с белками плазмы
| Тип нарушения | Уровень Т4 и Т3 в сыворотке крови | Процент СТ4 и СТ3 или ПТ3С | СТ4 и СТ3 или ИСТ4 и СТ3 |
| I. Первичное изменение ТСГ | |||
| Повышенная концентрация | повышается | снижается | Н |
| Сниженная концентрация | снижается | повышается | Н |
| II. Первичное нарушение функции щитовидной железы | |||
| Гипотиреоз | снижается | ||
| Гипертиреоз | повышается | повышается | повышается |
Обозначения: СТ4 — свободный Т4; СТ3- свободный Т3; ИСТ4 — индекс свободного Т4; ИСТ3-индекс свободного Т3; ПТ3С-поглощение Т3 смолой; ТСГ-тироксинсвязывающий глобулин.
| Повышенный уровень ТСГ | Сниженный уровень ТСГ |
| Беременность Неонатальный период Пероральные контрацептивы и другие источники эстрогенов Тамоксифен Инфекционный и хронический активный гепатит Билиарный цирроз Острая интермиттирующая порфирия Перфеназин Генетические причины Андрогенные и анаболические стероиды Большие дозы глюкокортикоидов Хронические заболевания печени Активная акромегалия Некроз Генетические причины Аспарагиназа | Андрогенные и анаболические стероиды Большие дозы глюкокортикоидов Хронические заболевания печени Активная акромегалия Некроз Генетические причины Аспарагиназа |
В первом случае ось щитовидная железа — гипофиз не нарушена, и гомеостатическая регуляция секреции тиреоидных гормонов сохраняется. В таких условиях нарушение взаимодействия обусловливается изменением связывания тиреоидных гормонов. Например, увеличение уровня ТСГ вначале снижает концентрацию свободного гормона и тем самым уменьшает его доступность для тканей. Затем общая концентрация гормона в сыворотке возрастает до тех пор, пока концентрация свободного гормона не восстановится до нормы. При этом доли свободных Т4 и Т3 снижаются. Увеличение общей концентрации гормона уравновешивает снижение доли свободной его формы и в результате абсолютная концентрация свободного гормона остается нормальной, что определяет и нормальный метаболический статус.
При снижении концентрации ТСГ происходят противоположные изменения. Состояния, сопровождающиеся первичными сдвигами в концентрации ТСГ, перечислены в таблице. Первичные нарушения связывания тиреотропных гормонов происходят при увеличении содержания в плазме и других связывающих белков, а также при появлении патологических связывающих белков. Эти вопросы обсуждаются ниже.
Во втором случае нарушение связывания тиреоидных гормонов обусловлено первичными изменениями их концентрации в крови, как это характерно для гипотиреоза или тиреотоксикоза. При этом нормальная гомеостатическая регуляция секреции тиреоидных гормонов теряется либо из-за нарушения самих регуляторных механизмов, либо потому, что интактные регуляторные механизмы оказываются неспособными преодолеть эффекты какой-либо патологии вне гомеостатической системы. В таких условиях концентрация ТСГ почти не меняется, а концентрация свободного гормона ока-зывается прямо пропорциональной его общей концентрации. Поскольку гомеостатиче-ские механизмы не могут восстановить нормальную концентрацию свободного гормона, первичные нарушения функции щитовидной железы сопровождаются постоянными измене-ниями концентрации общего и свободного гормона и, следовательно, изменениями ме-таболического статуса. При таких нарушениях доля свободного гормона меняется в том же направлении, что и поступление гормона в кровь.
Гормоны щитовидной железы ( T3, T4, кальцитонин): Метаболизм гормонов.
После своего проникновения в клетку Т4 и Т3 вступают в различные реакции, которые в конце концов приводят к их экскреции или инактивации. Метаболизм тиреоидных гормонов сводится главным образом к последовательному удалению каждого атома йода (монодейодирование) с образованием в конечном счете полностью лишенного йода тиронинового ядра.
Дейодированню подвергаются примерно 70% Т4 и Т3. В случае Т4 наибольшую важность имеет 5′-монодейодирование, которое приводит к образованию Т3 (Т3-неогенез). Поскольку около 30% Т4 превращается в Т3 и поскольку Т3 обладает примерно втрое большей метаболической активностью, чем Т4, практически весь метаболический эффект Т4 может быть отнесен на счет образующегося из него Т3. В нормальных условиях Т3-неогенез определяет примерно 80% присутствующего в крови Т3 и его общей продук-ции; остальное количество непосредственно секретируется щитовидной железой. По-этому патологические состояния и фармакологические средства, которые нарушают Т3-неогенез, понижают концентрацию Т3 в сыворотке.
Когда больные с гипофункцией щитовидной железы получают такие дозы синтетического Т4 (левотироксина), которые поддерживают его концентрацию в сыворотке на нормальном или слегка повышенном уровне, в крови создается нормальная или почти нормальная концентрация Т3.
Состояния, сопровождающиеся снижением периферической конверсии Т4 в Т3:
I. Физиологические:
Эмбриональный и ранний неонатальный период
Старческий возраст
II. Патологические:
Голодание
Нарушение питания
Системные заболевания
Физическая травма
Послеоперационный период
Фармакологические средства (пропилтиоурацил, дексаметазон, пропранолол, амиодарон)
Рентгеноконтрастные средства (ораграфин, телепак)
Положение о том, что щитовидная железа секретирует сравнительно небольшие количества Т3, неприменимо к состояниям, при которых имеет место автономная гиперфункция щитовидной железы, избыточная ее стимуляция ТТГ или сниженное содержание йода в ней. В таких условиях отношение Т3/Т4 в продуктах секреции щитовидной железы и в крови увеличивается.
Кроме того, при сниженной продукции Т4, как это наблюдается на ранних стадиях тиреоидной недостаточности или при дефиците йода, отношение концентраций Т3/Т4 в крови возрастает еще больше в ре-зультате срабатывания ауторегуляторного механизма, повышающего эффективность Т3-неогенеза.
Примерно 40% Т4 подвергается монодейодированию в положении 5 внутреннего кольца с образованием 3,3′, 5′-трийод-L-тиронина (реверсивный Т3, рТ3). Этот процесс определяет почти всю продукцию рТ3 в организме. Реверсивный Т3 практиче-ски не обладает метаболической активностью. Поэтому соотношение между процессами монодейодирования наружного и внутреннего колец детерминирует количество доступ-ного для тканей метаболически активного гормона. Факторы, нарушающие Т3-неогенез, почти всегда увеличивают концентрацию рТ3 в сыворотке.
Это увеличение связано не с повышенной продукцией рТ3 из Т4, а с торможением 5′-монодейодирования рТ3, в результате которого образуется 3,3′-дийодтиронин (3,3′-Т3). Иными словами, как снижение конверсии Т4 в Т3, так и снижение деградации рТ3 обусловлено избирательным нарушением 5′-монодейодирования.
Второй главный путь метаболизма Т4, Т3 и их метаболитов заключается в их конъюгировании в основном с глюкуронатом и сульфатом в печени. Эти конъюгаты либо подвергаются дейодированию на месте, либо выделяются в желчь, но размеры энтерогепатического кругооборота у человека неизвестны. Даже в лучшем случае про-исходит неполная реабсорбция и на долю экскреции Т4, Т3 и их йодсодержащих метаболитов с калом приходится примерно 20% общей элиминации Т4. Небольшая доля Т4 и Т3 (около 20%) подвергается окислительному дезаминированию и декарбоксилированию по боковой цепи аланина с образованием уксуснокислых аналогов — тетрайод- и трийодтироуксусных кислот (тетрак и триак соответственно).
В некоторых условиях изменения скоростей метаболического клиренса Т4 и Т3 определяются в основном сдвигами в накоплении и метаболизме гормонов. Фенобарбитал и фенитоин ускоряют метаболический клиренс тиреоидных гормонов, не увеличивая долю свободных гормонов в крови. Больше того, что касается фенитоина, то он снижает концентрацию общего и свободного Т4. Тем не менее нормальный метаболический статус поддерживается, вероятно, за счет повышения Т3-неогенеза.
Гормоны щитовидной железы ( T3, T4, кальцитонин): Действие гормонов.
Тиреоидные гормоны влияют на рост и созревание тканей, общие энергозатраты и кругооборот практически всех субстратов, витаминов и гор-монов, включая и сами тиреоидные гормоны. Первичные механизмы возникновения этих эффектов остаются неясными, но, по-видимому, гормоны действуют координирование на уровне клеточного ядра (изменяя экспрессию генома), на уровне митохондрий (влияя на окислительный обмен) и на уровне плазматической мембраны (регулируя поток субстратов и катионов в клетку и из нее).
Источник статьи: http://meduniver.com/Medical/Abdomen/44.html
Гормоны щитовидной железы
глава из книги «Анализ крови при болезнях щитовидной железы», А.В. Ушаков, 2016
Наконец, перед Вами оказался бланк с результатом анализа крови. Как же правильно понять лабораторные значения и не ошибиться?
Для освоения лабораторно-медицинского языка предлагаю, прежде всего, получить базовые знания. Важно узнать:
— какое конкретное действие оказывают гормоны щитовидной железы,
— где и как гормоны образуются, как они попадают в кровь,
— что влияет на образование гормонов,
— какие виды гормонов существуют, как они отражаются в лабораторных документах (данных анализа крови),
— какие показатели нужно определять всегда, а какие лишь в некоторых случаях.
Сразу предупрежу вас, читатель. Этот раздел может показаться сложным. Поэтому постарайтесь воспринимать его вдумчиво. При необходимости, перечитывать то, что с первого раза оказалось не совсем ясно. Если у вас получится усвоить знания этого раздела, то дальнейшее прочтение книги станет проще и интереснее.
В изучении науки понимания результатов анализа крови важны ваши старания. Приложенные усилия обязательно принесут вам существенную пользу.
Значение щитовидных гормонов в организме
Большинство клеток щитовидной железы образуют так называемые калоригенные гормоны (Т4 и Т3). Лишь малая часть специальных клеток железы выделяет гормон кальцитонин, участвующий в обмене кальция и фосфора вместе с паратгормоном, производимым околощитовидными железами.
Всё основное внимание в этой книге будет направлено на пояснение действия и особенностей калоригенных гормонов. Почему они так называются? Гормоны щитовидной железы (Т3 и Т4) участвуют в энергетическом обмене, который также именуется «основным обменом».
Энергию (в виде химических связей) человек получает с продуктами питания, а именно в виде углеводов и жиров (белки преимущественно используются в качестве «строительного» материала). Усвоение этой энергии клетками разных органов и систем оказывается возможно только с помощью щитовидных гормонов (Т3 и Т4).
Поэтому при увеличении потребления энергии (измеряется джоулями и калориями, отчего гормоны ― калоригенные) организм вынужденно стимулирует свою щитовидную железу, заставляя её производить и выделять в кровь больше своих гормонов. Такая стимуляция преимущественно исходит из двух мест: со стороны центров периферической нервной системы (прямыми нервными импульсами от нервных клеток к клеткам железы) и со стороны гипофиза (с помощью ТТГ).
Любые процессы в организме происходят с затратами энергии. Поэтому вместе с выделением энергии расходуются щитовидные гормоны.
Принципиальное устройство ткани щитовидной железы
Ткань щитовидной железы специально создана для производства важных организму гормонов. Вся структура железы состоит из бóльших и меньших сегментов. Самым малым типовым участком ЩЖ является фолликул, который и служит местом образования и хранения гормонов.
Подобно административному делению нашей страны, разграниченной на регионы, области, районы и населённые пункты (деревни, посёлки и города), щитовидная железа также состоит из крупных участков, включающих меньшие. В этом сравнении, населёнными пунктами, где, собственно, и происходят процессы производства гормонов, служат фолликулы. В щитовидной железе насчитывают несколько миллионов фолликулов. Вот как много «населённых пунктов» в огромной стране ― щитовидной железе. Что же такое фолликул, как он организован?
Фолликулы состоят из одного слоя клеток, окружающих по всему периметру некоторое количество белкового вещества, называемого коллоидом. Клетки щитовидной железы относятся к внутреннему эпителию. Эти клетки обладают способностью воспринимать из крови необходимые вещества, преобразовывать их и направлять обратно в кровь гормоны и продукты обмена.
Клетки обладают способностью захватывать частички коллоида, в котором хранятся гормоны, и передавать такой коллоид в кровь. Поскольку коллоид состоит из белка (тиреоглобулина), удерживающего гормоны, то в крови можно выявить соответствующее количество такого белка и гормонов щитовидной железы.
Каждый фолликул очень мал. Его размер в среднем почти в пять раз меньше миллиметра. Кроме того, встречаются микрофолликулы и макрофолликулы.
Микрофолликулы содержат мало коллоида. Часто они являются недавно сформировавшимися молодыми структурами. Им ещё предстоит «повзрослеть», увеличить функциональные возможности клеток, накопить коллоид и стать способными в достаточном количестве производить гормоны.
Макрофолликулы формируются при перенапряжении щитовидной железы. Они могут преобразовываться. Некоторые специалисты относят к макрофолликулам не крупные фолликулы, а объединения фолликулов: в случае индивидуальной предрасположенности, несколько фолликулов, соединённых между собой так называемыми окнами (между их коллоидным содержимым), объединяют свой коллоид и окружают клетками такое коллоидное «озерцо».
Образование гормонов происходит в клетках и коллоиде фолликула. От клеток в коллоид проходит своеобразный биохимический конвейер ― путь создания гормонов. Этот гормонообразующий конвейер направлен в обе стороны: из клетки в коллоид и из колоида в клетку.
Поступающие из крови во внутрь клеток вещества соединяются, окисляются и направляются из клетки в коллоид, где удерживаются белком (тиреоглобулином), составляющим основу коллоида. В коллоиде первичные структуры щитовидных гормонов, обозначаемые как Т1 (монойодтирозин) и Т2 (дийодтирозин), соединяясь с атомами йода и между собой, становятся готовыми гормонами ― Т3 и Т4. Гормон Т3 называется трийодтиронин, а Т4 ― тетрайодтиронин или короче ― тироксин.
К каждому фолликулу подходят нервные волокна и мелкие сосуды, привносящие продукты питания и вещества для производства гормонов и отводящие всё, что образуется в результате деятельности клеток. Фолликулы оплетены капиллярной сетью. Поверхности щитовидных клеток плотно контактируют со стенками капилляров.
Клетки фолликулов захватывают из капилляров (т.е. из крови) йод и аминокислоты, а из коллоида (внутри фолликулов), при потребности, ― готовые щитовидные гормоны, направляя их в сосудистое русло, к различным органам и тканям. Этот биохимический процесс регулируется и находится под прямым контролем периферической вегетативной нервной системы.
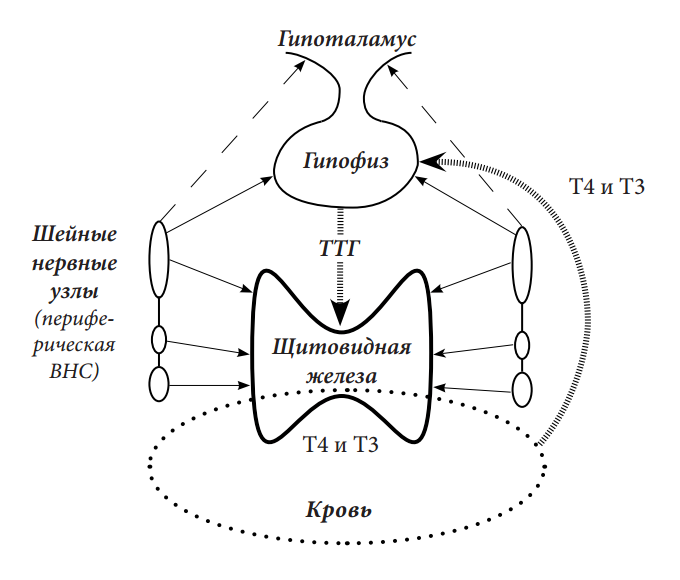
Рисунок 1. Схема регуляции деятельности щитовидной железы.
Периферическая вегетативная нервная система (ВНС), через нервные волокна от клеток в нервных центрах (в т.ч. в нервных узлах шеи), контролирует и регулирует деятельность щитовидной железы и гипофиза. Поэтому периферическая ВНС является основным источником управления деятельности щитовидной железы.
Выделяемые в кровь щитовидные гормоны (Т4 и Т3) по сосудистой сети достигают гипофиза. В зависимости от концентрации этих гормонов в крови, производится соответствующее количество ТТГ, с помощью которого оказывается стимуляция щитовидной железы (совместно с ВНС).
Периферическая ВНС может оказывать возбуждающее или тормозящее влияние на гипофиз.
Образование щитовидных гормонов
Для формирования щитовидных гормонов необходим йод. Этот элемент поступает в организм с водой и пищей. Взрослому человеку обычно нужно около 150 мкг йода в сутки. При увеличении потребности организма в гормонах Т3 и Т4, соответственно увеличивается количество усваиваемого йода ― от 200 мкг в сутки и более.
Лишний йод выводится преимущественно с мочой (почти 97%) и весьма мало через кишечник, куда йод попадает с желчью из печени (при фильтрации крови в ней). Увеличение потребления йода щитовидной железой из крови приводит к уменьшению выделения йода с мочой. Поэтому оценка количества йода в моче может помочь в диагностике и выборе лечения.
В нашей Клинике рекомендуем определение концентрации йода в моче при гипотиреозе и некоторых вариантах эутиреоза. Исследование йода мочи таких пациентов показало, что дефицит йода у жителей России наблюдается относительно редко, почти в 10-15% случаев. Совместная интерпретация гормональных показателей крови и йода мочи точнее раскрывает индивидуальные особенности болезни.
Щитовидная железа очень активно захватывает йод. Поэтому концентрация йода в ткани железы в 20-40 раз больше, чем в крови. Регуляция потребления йода щитовидной железой регулируется нервной системой, ТТГ (образуется гипофизом), тиреоглобулином и йодом. Нервная система и ТТГ способны непосредственно усиливать напряжение щитовидной железы и скорость потребления йода. Накопление тиреоглобулина в клетках железы тормозит захват йода. Избыточное поступление йода в кровь уменьшает образование гормонов (этот феномен несколько десятилетий назад применяли для подавления деятельности щитовидной железы при тиреотоксикозе).
Поступающий в фолликулы йод первоначально окисляется благодаря специальному ферменту тиреопероксидазе (ТПО) и перекиси водорода, которые создаются в клетках специально для окисления йода. Затем ион йода (I+) соединяется с тирозином и образует монойодтирозин (МИТ или Т1) ― первый предшественник щитовидных гормонов. Два иона йода формируют дийодтирозин (ДИТ или Т2).
Лишнее количество перекиси водорода нейтрализуется для предотвращения повреждения клеток щитовидной железы. Такое «антиоксидазное» действие осуществляется специальными ферментами и соединениями белка с селеном.
Это первый участок биохимических процессов, в котором выявлена важная роль селена. Другие участки находятся в печени, почках и некоторых других тканях, где селен применяется для отщепления иона йода от Т4 (тироксина) и создания главного потребляемого гормона ― Т3 (трийодтиронина). Вот почему так важно оценивать достаточность селена в организме и своевременно пополнять его (но следует не переусердствовать в этом).
Образование МИТ и ДИТ происходит с участием тиреоглобулина, специального белка, находящегося в коллоиде фолликулов. Этот процесс замечен исследователями на границе клетки и коллоида.
Предшественники щитовидных гормонов (МИТ и ДИТ) в последующем объединяются, создавая гормоны ― Т4 и Т3. Соединение двух молекул ДИТ (т.е. Т2+Т2) образует гормон Т4, а при взаимодействии МИТ и ДИТ (т.е. Т1+Т2) ― гормон Т3. В этот процесс также вовлечён фермент ТПО (тиреопероксидаза). Таким образом, ТПО принимает участие во всех ключевых этапах гормонообразующего конвейера щитовидной железы.
Тиреоглобулин в коллоиде фолликулов удерживает МИТ, ДИТ, Т3 и Т4. Это обстоятельство необходимо для создания гормонов. Вместе с тем, предгормоны и гормоны, соединённые с тиреоглобулином, служат важным гормональным запасом. Этот источник способен некоторое время обеспечивать организм гормонами даже в случае прекращения поступления в организм йода. Но гормональный склад истощаем и потому требует регулярного пополнения. Относительно активное разрушение щитовидной железы, встречающееся при остром тиреоидите, может способствовать поступлению в кровь очень большого количества щитовидных гормонов. В таком случае анализ крови покажет признаки гипертиреоза. Но этот избыток гормонов не стойкий. Относительно быстро (по мере прекращения воспаления и поступления гормонов в кровь из разрушенных фолликулов) выраженность гипертиреоза уменьшается, и гипертиреоз вновь переходит в эутиреоз.
Особенности щитовидного гормонального обмена
Постарайтесь внимательно прочесть и понять содержание этого подраздела. Следующие сведения очень важны для оценки анализа крови. Эти знания послужат ключом к пониманию данных анализа.
Врачи называют гормоны «свободными», «связанными» и «общими». Такие обозначения могут показаться странными. Тем не менее, именно они служат смысловой опорой в понимании результатов анализа крови. Что же подразумевается под этими терминами?
Напомню, что гормоны щитовидной железы представляют собой органические молекулы. Из щитовидной железы гормоны (Т4 и Т3) поступают в кровь в свободном виде, т.е. не соединённые с белками или с чем бы то ни было.
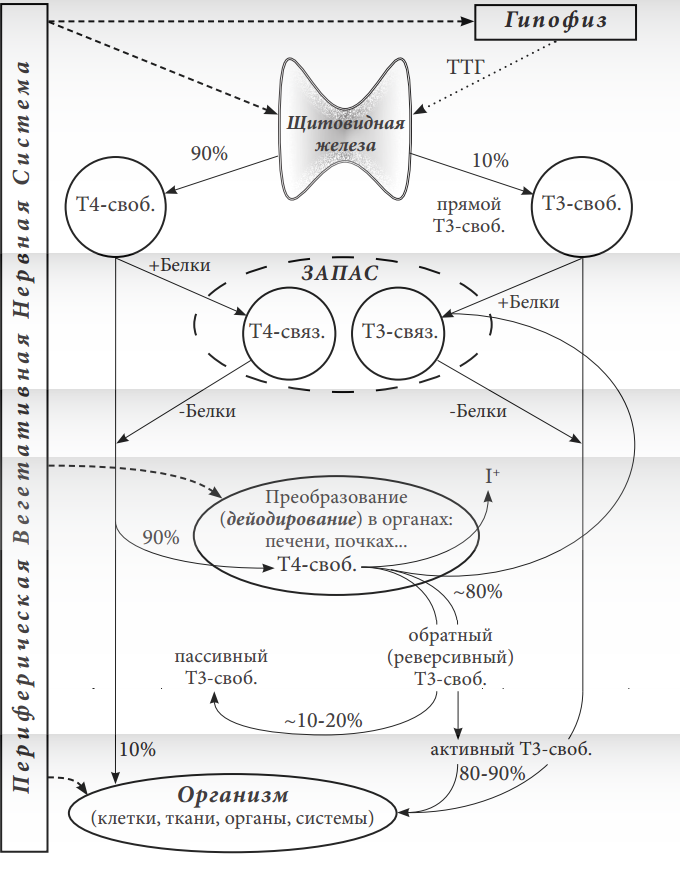
Рисунок 2. Схема обмена гормонов щитовидной железы (Т4 и Т3) с пояснениями.
1 уровень. Образование и выделение свободных Т4 и Т3 из щитовидной железы в кровь. В образовании гормонов принимает участие селен. Этот процесс контролируется и регулируется периферической ВНС и гипофизом.
2 уровень. Большая часть свободных Т4 и Т3 соединяется в крови с белковыми молекулами. В результате образуются связанные Т4 и Т3. Такие комплексы гормонов с белками неактивны и представляют запас.
3 уровень. При необходимости, гормоны Т4 и Т3 отсоединяются от белков и вновь становятся свободными.
4 уровень. Из 90% гормона Т4-свободного путём отсоединения атома йода образуется Т3-свободный (отделяющиеся ионы йода попадают с током крови в щитовидную железу, где вновь используются). В этом процессе дейодирования Т4св. принимают участие ферменты, содержащие селен. Регуляция дейодирования происходит под влиянием периферической ВНС.
5 уровень. Организм потребляет около 10% Т4св. и почти 80% активного Т3св. Это усвоение гормонов регулируется периферической ВНС. Пропорции гормонов колеблются в зависимости от компенсаторного состояния организма.
Из всего количества щитовидных гормонов 99,96% Т4 и 99,6% Т3 становятся связанными с белковыми молекулами. Соответственно, лишь 0,04% Т4 и 0,4% Т3 остаются свободными. Только свободные гормоны оказывают биологическое действие.
Представьте, что этого «малого» количества гормонов (от общего их количества в крови) достаточно для обеспечения энергетического обмена в организме. Этот факт сообщает нам о значительной калоригенной возможности щитовидных гормонов и их больших запасах!
Связанные гормоны не активны. Белки, соединяясь с гормонами, таким образом резервируют главный продукт щитовидной железы, постоянно поступающий в кровь. Этим предотвращается как потеря гормонов, так и перенасыщение гормонами организма.
Выявлено несколько видов белков, связывающих щитовидные гормоны. Одни виды белков создают более прочные соединения, а другие, напротив, способны быстрее высвободить гормоны.
Связанные щитовидные гормоны представляют запас организма, который при необходимости может быть использован. Разная степень «прочности» соединения белков крови с гормонами позволяет быстро обеспечивать организм щитовидными гормонами в нужном количестве. Такое срочное насыщение гормонами особенно важно при острых заболеваниях и истощающих перегрузках.
Лаборатории, к сожалению, не ориентированы на выявление связанных фракций щитовидных гормонов. Лаборатории анализируют величины свободных и общих гормонов. В общих Т4 и Т3 заключены соответствующие им свободные и связанные формы гормонов.
Обратите внимание на то, что свободной фракции Т3 (Т3св.) в крови в 10 раз больше, чем свободной фракции Т4 (Т4св.). Это обстоятельство указывает на ведущую роль в обмене гормона Т3. Организм стремится поддерживать достаточное количество и оптимальную концентрацию этого гормона в крови.
По мнению разных исследователей, из щитовидной железы в кровь поступает около 80-90% Т4 и, соответственно, ― 10-20% Т3 (не путайте проценты поступающих в кровь гормонов с процентами связываемых белками гормонов). Независимо от того, что щитовидная железа производит значительно больше Т4, организм (все его ткани и органы) потребляет главным образом Т3 (рис. 2).
Дело в том, что гормон Т4 в 4-5 раз менее активен по сравнению с гормоном Т3. Это следует понимать так, что Т4 медленнее и меньше по сравнению с Т3 позволяет «усваивать» энергию. Т4 преимущественно является «промежуточной» химической структурой.
Из всего произведенного щитовидной железой количества Т4 организм потребляет лишь 10% (рис. 2). Остальные 90% выделенного щитовидной железой Т4 преобразуются в Т3. Поэтому Т4 можно рассматривать как полуфабрикат, из которого создаётся главный гормон ― Т3. Природа организма специально производит больше менее активного гормона. Иначе мог бы возникнуть риск пресыщения организма энергией. Рациональнее по мере надобности создавать основной гормон (Т3) из прогормона (Т4).

Цитаты из книги
М.И. Балаболкин, Е.М. Клебанова, В.М. Креминская «Фундаментальная и Клиническая Тироидология». Руководство для врачей. М.: Медицина, 2007, 816 с.
В следующем изложении авторы под Т3 и Т4 представляют свободные формы щитовидных гормонов.
Стр. 121: «Наибольшее биологическое действие имеет Т3, который активнее Т4 в 4-5 раз. В течение длительного времени считалось, что Т4 и Т3 в равной степени принимают участие в обмене веществ. Однако было показано, что на периферии Т4 конвертируется (переходит) в Т3 и биологическое действие тиреоидных гормонов более чем на 90-92% осуществляется за счёт Т3. Применение радиоиммунологического метода для определения концентрации Т3 в сыворотке крови позволило установить, что около 80% циркулирующего в крови Т3 является производным от Т4 в следствие его периферического монодейодирования, и только 20% Т3 сыворотки непосредственно образуется в щитовидной железе».
Стр. 122: «… почти 100% действия тиреоидных гормонов осуществляется Т3, а не Т4…».
Стр. 134: «Основная роль тироксина в организме заключается в том, что он является своего рода источником или, правильнее, прогормоном, для Т3, который и оказывает биологическое действие в тканях организма».
Преобразование Т4 в Т3 происходит, главным образом, в печени, но также возможно в почках и некоторых других структурах. Этот процесс осуществляется путём отделения из молекулы Т4 одного атома йода (четвёрка в формуле Т4 означает количество атомов йода). Название процесса ― дейодирование, т.е. отщепление йода.
Отделение атома йода из молекулы Т4 происходит с помощью специального фермента ― дейодиназы. Различают три вида дейодиназ, характеризующихся разным местом нахождения и особенностями своего влияния.
В процессе преобразования Т4 в Т3 участвует селен. Этот элемент входит в состав дейодиназ, которые представляют собой соединение белка с селеном. Поэтому достаточность селена ― это важное условие для полноценности дейодирования Т4 (т.е. отсоединения от Т4 одного атома йода) и образования наиболее важного для организма гормона ― Т3.

Цитаты из книги
Г.М. Кроненберг, Ш. Мелмед, К.С. Полонски, П.Р. Ларсен «Заболевания щитовидной железы». Пер. с англ. под ред. И.И. Дедова, Г.А. Мельниченко. М.: ООО «Рид Элсивер», 2010, 392 с.
Авторы сообщают, что в организме из Т4св. образуется 80% активного гормона Т3св., путём отсоединения от него одного атома йода (с помощью специальных ферментов ― дейодиназ). Лишь малая часть гормона Т3 неактивна.
Стр. 37: «Нужно отметить, что тканями захватываются, действуют на них, подвергаются дейодированию и инактивации только свободные тиреоидные гормоны. Связанная же их форма служит резервуаром».
Стр. 40: «Наиболее значимый путь метаболизма Т4 ― его монодейодирование по наружному кольцу в 5ʾ-положении. Результатом может быть продукция активной формы тиреоидных гормонов, Т3. Эта реакция катализируется дейодиназами 1-го и 2-го типов (D1 и D2) и служит источником примерно 80% всего Т3, циркулирующего в крови человека».
Авторы указывают на ведущее участие периферической вегетативной нервной системы (её симпатической части) в регуляции образования Т3св. из Т4св., путём влияния на активность дейодиназы D2 [уточнение: периферическая ВНС способна реагировать на любые влияния (в т.ч. холод) и при любом виде стресса стимулировать обмен щитовидных гормонов (и напряжение ЩЖ)].
Стр. 45: «Поскольку экспрессия кодирующего D2 гена Din2 возрастает под влиянием цАМФ, в бурой жировой ткани при холодовой стимуляции симпатических нервных волокон быстро нарастают активность D2 и продукция Т3».
Т3св. ― важный и обязательный показатель!
Т3-свободный является сущностно важным гормональным показателем щитовидного обмена! Только Т3св. преимущественно потребляется организмом и лучше оказывает энергообразующее действие, чем Т4св.
Т3св. современные специалисты предлагают определять вместе с ТТГ и Т4св. лишь при тиреотоксикозе, но абсолютно игнорируют при всех прочих состояниях гормонального обмена.
Более того, авторитетные эндокринологи России определение Т3св. в случае гипотиреоза относят к ошибкам (они не видят смысла в их оценке), а некоторые лаборатории (таких мало) вообще отказывают пациентам и врачам в определении этого показателя. Чем же такие специалисты объясняют своё «вето» на Т3св.?
Усматривают две основные причины такого игнорирования Т3св. Поскольку количество Т3св. организм стремиться сохранить в достаточном (усреднённом) количестве, то более часто встречающиеся «нормальные» величины Т3св. по сравнению с Т4св. служат аргументом для исключения показателя. Другая причина вызвана неким знанием о существовании не только активно действующей формы Т3св., но и нейтральной формы Т3св.
Оказывается, в зависимости от места расположения (прикрепления) атомов йода в молекуле гормона, существует несколько разновидностей Т3. В одном случае молекула Т3 активна, но в другом ― пассивна.
Щитовидная железа производит и выделяет так называемый прямой гормон Т3. Обычно полагают, что это активный гормон. Его малое количество недостаточно для полноценной жизни. Дополнительно, путём отсоединения йода от Т4, в нужном количестве образуется гормон Т3, называемый реверсивным (или обратным).
Структура реверсивного Т3 может быть идентична прямому Т3 или отличаться другим расположением атомов йода в молекуле. В первом случае Т3 будет оказывать полноценное действие в организме, во втором случае гормон Т3 будет нейтральным.
Интересно, что в любом конкретном случае невозможно оценить по анализу крови количество активного и пассивного Т3св. Поэтому не спешите заказывать в лаборатории определение таких точных показателей. Неизвестно, как ведет себя конкретный организм при эутиреозе, гипотиреозе и гипертиреозе в отношении формирования вариантов Т3св. Исследователям удаётся определять лишь реверсивный Т3св. (большинство лабораторий также не определяют этот показатель), который может быть как активным, так и пассивным. В результате весь Т3св. необоснованно воспринимается как малоинформативный из-за «неизвестного» соотношения в нём активных и пассивных форм гормона Т3.
Противники оценки Т3св. мотивируют своё мнение двумя представленными причинами. Вместе с тем, они указывают на расход средств, затрачиваемых на определение «лишнего» показателя. Так ли всё на самом деле?
История науки и в т.ч. медицины показывает, что во все времена и относительно часто возникают различные гипотезы (предположения), которые восполняют «пробелы» в реальных знаниях. По мере накопления и понимания фактов, противоречащих используемой гипотезе, формируется более достоверное суждение, точнее соответствующее законам мира. Оценке лучшего знания помогает практика, подтверждая и уточняя особенности нового понимания в каждом исследуемом случае.
Главное в совершенствовании нашего мировоззрения заключается в умении видеть факты и понимать их. Но не все люди способны замечать подсказки природы и совершать открытия. Леонардо да Винчи приписывают такие мудрые строки: «Одни люди видят, другие ― видят, когда им поясняют, третьи ― не видят». К этому можно добавить нежелание познавать и совершенствоваться, а также зависимость от авторитетного мнения людей, занимающих должности и увенчанных регалиями…
В течение последних 12 лет мне почти каждый будний день удавалось многократно (на примерах разных пациентов) отслеживать и внимательно анализировать изменения Т3св. и других гормональных показателей. Я стремился понять особенности развития болезни при разных заболеваниях. Каждый (!) случай болезни подвергал тщательному анализу и делал выводы, почему показатели крови ведут себя именно так, а не иначе. Обязательно совмещал изучение данных анализа крови с другими исследованиями (сам проводил УЗИ, высматривая характер процессов в реальности, изучал данные сцинтиграфии, термографии и прочее). Для лучшего понимания, получал недостающие знания из книг и статей: о закономерностях развития болезней (из общей патологии человека), о процессах в щитовидной железе и за её пределами (эндокринологической литературе), о влиянии нервной системы, роли иммунной системы и т.д. (вместе с тем, за годы в нашей Клинике выросла целая научно-медицинская библиотека!).
В результате, пришёл к выводу о значительной роли гормона Т3св., о существовании искажений в понимании участия Т3св. в обмене и обязательности оценки этого показателя при анализе крови. Это решение не было скорым. Пришлось потратить годы.
Как и многие специалисты, изначально исходил из имеющихся авторитетных рекомендаций и определял совместно только два гормона ― ТТГ и Т4св. Интересующиеся могут увидеть такую мою позицию в первом издании книги для пациентов «Восстановление щитовидной железы», изданной в 2008 г. Но уже во втором издании этой книги (улучшенном и исправленном) в 2011 г. читатель сможет обнаружить полноценную рекомендацию от нашей Клиники ― всегда, как минимум, совместно определять три показателя: ТТГ, Т4св. и Т3св.
Что же думают по этому поводу авторитеты? Важен ли показатель Т3св.? Оказывается, тоже важен. Действительное знание пробивается через искажения, мифы, гипотезы и авторитетные «запреты».
Все (в т.ч. авторитетные) современные клинические научные издания содержат рекомендацию определять Т3св. при гипертиреозе (тиреотоксикозе) вместе с ТТГ и Т4св.! Почему? Потому что при гипертиреозе Т3св. не «удерживается» организмом в средних значениях нормы и стремится к увеличению. Именно избыток Т3св. является ведущим при тиреотоксикозе (гипертиреозе). Как это можно не заметить и пропустить? В случае гипертиреоза (тиреотоксикоза) никто из специалистов не позволяет себе «критиковать» Т3св. ― обвинять его в недемонстративности, ложности, неполноценности и пр. Более того, выделяют даже так называемый «Т3-свободный тиреотоксикоз» (когда Т4св. находится в норме).
И это не всё. Специалисты «создали» особое «заболевание», имеющее разные названия, одно из которых ― «Синдром низкого Т3св.». Такое уменьшение Т3св. сообщает об увеличении потребности организма в щитовидных гормонах (этот случай представлен в этой книге в разделе об Эутиреозе). При таком синдроме ТТГ может находиться в норме (в эутиреозе).
Получается, что при гипертиреозе и эутиреозе Т3св. нужно контролировать! Чем же оказался неугоден Т3св. при гипотиреозе? Почему этот показатель не нужно оценивать при увеличении ТТГ (признаке гипотиреоза)? В чём сущность этого странного противоречия (при эутиреозе и гипертиреозе Т3св. нужно определять, а при гипотиреозе ― нет)?
Наблюдения в нашей Клинике выявили закономерные колебания Т3св. не только при гипертиреозе и эутиреозе. Совершенно аналогичные изменения Т3св. постоянно встречаются и при гипотиреозе! Может быть, это случайность? Но многочисленность наблюдений и оценка изменений гормонального обмена у таких пациентов на протяжении разных по длительности периодов позволят утверждать, что приписываемые гормону Т3св. малоинформативные свойства не подтверждаются. Более того, по мнению нашей Клиники, Т3св. является ключевым по сравнению с прочими показателями гормонального обмена щитовидной железы.
Различные исследования, при которых также анализировался показатель Т3св., подтверждают его важное значение. Можно сказать, что природная закономерность роли Т3св. всегда в разной степени проявляет себя. Это знание не скрыто. Оно явно.
Во время международной конференции по раку щитовидной железы в мае 2013 года один из иностранных докладчиков обратил внимание на связь между достаточным количеством Т3св. (по данным анализа крови) и стрессовой устойчивостью. Мой уточняющий вопрос о значении Т3св. и обязательности оценки этого показателя остался без ответа.
Иногда в моей практике встречаются пациенты с очень напряжённой формой гипотиреоза. У них значительно более нормы увеличено количество ТТГ (признак стимуляции щитовидной железы) и уменьшено Т4св. (менее нормы). При этом они не применяют гормональные средства, почти не предъявляют жалоб и ведут обычный образ жизни, не подозревая об изменении гормонального обмена. Почему при гипотиреозе с сильно выраженным недостатком Т4св. такие люди чувствуют себя замечательно?
Ответ прост: потому что у таких пациентов полноценно количество Т3св.! У этих пациентов оптимальное значение Т3св. наблюдается за счёт очень интенсивного преобразования Т4св. в Т3св. При этом их щитовидная железа очень интенсивно производит гормоны (побольше Т4 и поменьше Т3), а заставляет перенапрягаться железу ТТГ (вместе с периферической нервной системой).
Если и далее гипотетически предполагать, что Т3св. неактивен (из-за иного расположения атома йода в молекуле), то почему сохраняется хорошие самочувствие и работоспособность пациента при увеличенном ТТГ и уменьшенном Т4св.? Такая ситуация возможна только в одном случае ― при абсолютной полноценности Т3св.!
При почти одних и тех же значениях ТТГ или сочетании ТТГ с Т4св. могут определяться разные количества Т3св. Важно выявить индивидуальные особенности гормонального обмена. Это поможет оценить и понять реальное состояние организма (включая его щитовидную железу и всё, что к ней прямо относится), предвидеть развитие процесса и выбрать более верное лечебное направление. Но как всё это можно сделать вслепую, исключая Т3св., Т4св. и ориентируясь только на ТТГ, как это предлагают и делают многие мои коллеги?!
Т3св. вместе с ТТГ и Т4св. служат обязательными показателями в понимании клинической ситуации. Без знаний о значении Т3св. специалист будет подобен детям капитана Гранта и их спутникам, тщетно искавшим отца по недостаточным данным (фото). Диагностика по ТТГ и Т4св. также будет соответствовать двухмерной ориентации в трёхмерном мире.
Для усвоения знаний первого раздела книги приведу пример. Думаю, вам будет понятен мой ответ пациентке на Форуме сайта нашей Клиники:
Ольга (Шадринск) — 21 октября 2015:
«Помогите пожалуйста, от приема Л-тироксина начинаются настоящие отеки, несколько раз лежала с диагнозом отек Квинке. Без приема гормонов отеки и крапивница прекращаются, но показатели ТТГ―13, Т4―11, а месяц назад Т4 был 16, а ТТГ―17, Подскажите как быть дальше?».
1. Главный потребляемый гормон ― это Т3св. Главный производимый гормон ― Т4, из которого потом образуется Т3, который в качестве Т3св. потребляется. Поэтому ВСЕГДА вместе с ТТГ и Т4св. нужно определять Т3св.
2. Уменьшение Т4 связано с увеличением потребления Т3св. и соответствующим усилением преобразования Т4 в Т3, в связи с чем Т4св. становится меньше.
3. ТТГ ― гормон гипофиза, заставляющий щитовидную железу создавать Т4 и Т3. Значения ТТГ 13 и 17 не очень сильно отличаются. Более того, ТТГ постоянно колеблется в некотором пределе, в зависимости от условий (они меняются, и приходится по-разному тратить энергию и энергетические гормоны (Т3 и Т4) в свободном виде).
4. При достаточности Т3св. можно думать о достаточности компенсаторных сил щитовидной железы, возможности некоторое время обойтись без дополнения гормонами извне (с препаратом).
5. Важно оценить с помощью УЗИ состояние щитовидной железы (в первую очередь, количество полноценной ткани и сосудистую активность).
6. Независимо от применения помощи гормонами важно влиять на основу болезни, устраняя необходимость тратить больше, чем обычно, гормонов. Оценить состояние железы, гормональный обмен и прочее, включая сущность болезни, должен врач.
Источник статьи: http://dr-md.ru/thyroid-hormone.html
Гормоны тиреоидной функции
Тиреотропный гормон
Строение
Представляет собой гликопротеин с молекулярной массой 30 кДа, состоит из двух субъединиц α- и β, α-субъединица схожа с таковой гонадотропных гормонов, β-субъединица специфична для ТТГ.
Синтез
Осуществляется в базофильных тиреотрофах гипофиза.
Регуляция синтеза и секреции
Активируют : тиреолиберин, охлаждение (закаливание, обливание холодной водой); также усиливается в вечернее время суток.
Уменьшают : соматостатин, кортизол, тироксин и трийодтиронин (по механизму обратной отрицательной связи).
Механизм действия
Аденилатциклазный, связанный с ингибированием GI-белка и накоплением цАМФ, и кальций-фосфолипидный механизм с образованием инозитол-трифосфата, диацилглицерола и комплекса кальций-кальмодулин.
Мишени и эффекты
Тиреотропный гормон в щитовидной железе:
1. Обеспечивает жизнедеятельность щитовидной железы
- стимулирует углеводный обмен (гликолиз, ПФП), синтез гетерополисахаридов,
- повышает синтез белков, фосфолипидов и нуклеиновых кислот,
- стимулирует васкуляризацию щитовидной железы,
- стимулирует рост и пролиферацию тиреоидных клеток,
2. Стимулирует гормональную активность щитовидной железы
- активирует натрий-йодидный транспортер на базолатеральной мембране тиреоцита, что повышает захват йода,
- активирует анион-транспортный белок на апикальной мембране тиреоцита, что увеличивает переход йода в фооликулярное пространство,
- активирует все стадии образования трийодтиронина и тироксина, в том числе увеличивает экспрессию гена тиреопероксидазы.
Патология
При уменьшении выработки изменяется масса тела, повышается утомляемость, возникают симптомы гипотиреоза (см ниже).
Йодтиронины
Строение
К гормонам самой щитовидной железы относятся тироксин и трийодтиронин, которые представляют собой йодированные производные аминокислоты тирозина.

Строение гормонов щитовидной железы
Синтез
Осуществляется в фолликулярных клетках щитовидной железы. Йодиды поступают через базолатеральную мембрану в фолликулярную клетку симпортом с ионами Na + , градиент последних создается Na + ,K + -АТФазой, далее перемещаются к апикальной мембране и выходят из клетки в фолликулярное пространство при участии анион-транспортного белка.
На апикальной мембране клеток селен-зависимая гемсодержащая тиреопероксидаза :
- йодирует остатки тирозина в тиреоглобулине с образованием моно- и дийодпроизводных (МИТ, ДИТ) тирозина,
- конденсирует часть МИТ и ДИТ до йодтиронинов, при этом доля трийодтиронина (Т3) и тетрайодтиронина (тироксин, Т4) составляет около 30% от всех йодпроизводных.
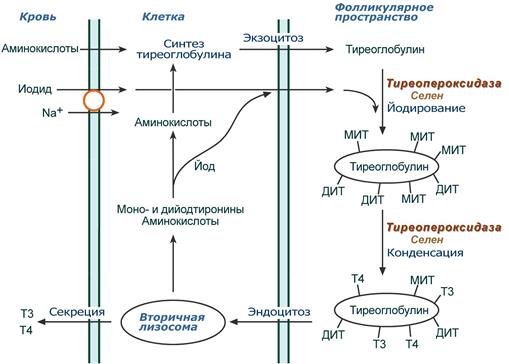
Схема реакций синтеза тиреоидных гормонов
Йодированный тиреоглобулин хранится в виде коллоида в просвете фолликула, при тиреотропной стимуляции пиноцитируется фолликулярными клетками, сливается с лизосомами и гидролизуется. Далее три- и тетрайодтиронин секретируются в кровь. В крови гормоны транспортируются специфическим глобулином, а также альбумином.
Неиспользованные моно- и дийодтирозины удерживаются в клетке и дейодируются йодтирозиндегалогеназой. Высвобожденный йодид вновь используется для синтеза гормонов щитовидной железы.
Регуляция синтеза и секреции
Активируют: тиреотропин на этапах поглощения йода, синтеза тиреоглобулина, эндоцитоза и секреции Т3 и Т4 в кровь.
Уменьшают: тироксин и трийодтиронин (по механизму обратной отрицательной связи).
Синтез гормонов также подавляют стрессы, инфекции, травмы, высокие концентрации йода (бесконтрольный прием препаратов KJ), соединения фтора, токсины (пестициды, кадмий, свинец, ртуть).
Механизм действия
Мишени и эффекты
Рецепторы к йодтиронинам имеют все ткани организма. В клетках-мишенях, особенно в печени, тироксин дейодируется и активной формой является трийодтиронин (3,5,3′-производное).
Деактивация тироксина в неактивный 3,3′,5′-трийодтиронин (reverse T3, rT3) происходит при участии деиодиназы (тип 3). Его увеличивают стресс, травмы, низкокалорийная диета. воспалительные процессы (цитокины), инфекции, дисфункция печени и почек, токсины и некоторые лекарства.
Превращение тироксина в активный 3,5,3′-трийодтиронин (деиодиназа 2) нуждается в ионах цинка и селена. Эта реакция ослаблена у плода, новорожденных и престарелых.
Тиреоидные гормоны увеличивают скорость базального метаболизма. Главным эффектом является повышение активности Na + ,K + -АТФазы , что приводит к быстрому расходованию АТФ и по механизму дыхательного контроля запускает катаболизм углеводов и липидов. В митохондриях увеличивается количество АТФ/АДФ-транслоказы и потребление кислорода. Сопутствующим эффектом усиления катаболизма является термогенез (наработка тепла).
Белковый обмен : Усиливает транспорт аминокислот в клетки. Активирует синтез дифференцировочных белков в ЦНС, гонадах, костной ткани и обусловливает развитие этих тканей.
У детей действие тиреоидных гормонов в целом анаболическое, т.к. трийодтиронин усиливает выделение соматолиберина, что стимулирует секрецию гормона роста. Одновременно он синергичен другим метаболическим эффектам СТГ, что и является причиной низкорослости при гипотиреозе.
У взрослых действие тиреоидных гормонов, в основном, катаболическое.
Углеводный обмен : Увеличивает гликогенолиз и аэробное окисление глюкозы.
Липидный обмен : Стимулирует липолиз, β-окисление жирных кислот, подавляет стероидогенез.
Нуклеиновый обмен : Активирует начальные стадии синтеза пуринов и синтеза пиримидинов, стимулирует дифференцировочный синтез РНК и ДНК.
- в надпочечниках подавляет синтез катехоламинов, хотя в целом чувствительность тканей к адреналину повышается.
Инактивация тиреоидных гормонов
Деактивация тиреоидных гормонов происходит в тканях-мишенях при действии дейодиназ, последовательно удаляющих от молекулы йод. Далее реакции катаболизма включают дезаминирование или декарбоксилирование боковой цепи или расщепление эфирной связи с образованием неактивных соединений. В печени дейодированные метаболиты связываются с глюкуроновой или серной кислотой и удаляются с желчью.
Патология
Гипофункция
Причина. Развивается при снижении синтеза тиреоидных гормонов в результате недостаточной стимуляции со стороны гипофиза и/или гипоталамуса, при заболевании самой железы, при нехватке необходимых веществ (аминокислоты, железо, йод, селен). Очень часто причиной выраженного гипотиреоза является болезнь Хашимото , при которой вырабатываются блокирующие антиантитела к рецепторам .
Клиническая картина
Симптомами субклинического гипотиреоза, зачастую неспецифическими, могут быть
- отечность лица, сухость кожи и волос, ломкость ногтей, увеличение массы тела, бледность, скованность мышц,
- галакторея у женщин в конце цикла,
- брадикардия, понижение систолического давления,
- психическая инертность, депрессия, апатия, вялость, сонливость, утомляемость, запоры,
- снижение утренней температуры тела до 36,0°-35,5°С и ниже.
У подростков отмечается отставание в физическом развитии, позднее половое созревание, функциональная дебильность, т.е. замедление мышления, снижение успеваемости в школе, неспособность к творческой деятельности, утрата чувства юмора.
При наличии выраженного гипотиреоза:
- у плодов, новорожденных и детей младшего возраста развивается кретинизм .
- у взрослых отмечается микседема , у женщин – бесплодие и галакторея (см регуляция синтеза и секреции пролактина),
- у обоих полов – деменция, психоз.
При лечении используют заместительную терапию левотироксином (L-тироксин).
Гиперфункция
Причина. Большинство случаев выраженного гипертиреоза вызвано наличием активирующих антиантител к рецепторам . В этом случае заболевание носит название болезнь фон Базедова (в отечественной и европейской литературе) или болезнь Грейвса (в американской литературе).
Клиническая картина.
Симптомами субклинического гипертиреоза являются
- трудность засыпания, эмоциональная лабильность и нервозность (плаксивость),
- выпадение волос, сухие ногти,
- неизменность веса на фоне повышения аппетита,
- тахикардия, мышечная слабость, потливость, влажные ладони.
При более выраженной форме отмечается субфебрильная температура (до 37,5°С), особенно к вечеру, нервное возбуждение, экзофтальм, тремор, диарея, похудание, у женщин бесплодие и скудость месячных.
При лечении используют тиреостатические препараты, которые ингибируют тиреопероксидазу (мерказолил, пропилтиоурацил), радиойодтерапию и хирургическое иссечение участка железы.
Источник статьи: http://biokhimija.ru/gormony/tiroksin.html