Инсулин — самый молодой гормон
Строение
Инсулин представляет собой белок, состоящий из двух пептидных цепей А (21 аминокислота) и В (30 аминокислот), связанных между собой дисульфидными мостиками. Всего в зрелом инсулине человека присутствует 51 аминокислота и его молекулярная масса равна 5,7 кДа.
Синтез
Инсулин синтезируется в β-клетках поджелудочной железы в виде препроинсулина, на N-конце которого находится концевая сигнальная последовательность из 23 аминокислот, служащая проводником всей молекулы в полость эндоплазматической сети. Здесь концевая последовательность сразу отщепляется и проинсулин транспортируется в аппарат Гольджи.
На данном этапе в молекуле проинсулина присутствуют А-цепь, В-цепь и С-пептид (англ. connecting – связующий). В аппарате Гольджи проинсулин упаковывается в секреторные гранулы вместе с ферментами, необходимыми для «созревания» гормона . По мере перемещения гранул к плазматической мембране образуются дисульфидные мостики, вырезается связующий С-пептид (31 аминокислота) и формируется готовая молекула инсулина. В готовых гранулах инсулин находится в кристаллическом состоянии в виде гексамера, образуемого с участием двух ионов Zn 2+ .
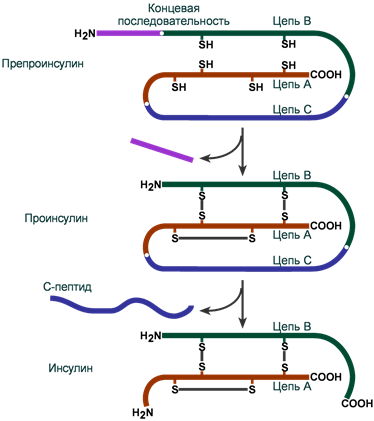
Схема синтеза инсулина
Регуляция синтеза и секреции
Секреция инсулина происходит постоянно, и около 50% инсулина, высвобождаемого из β-клеток, никак не связано с приемом пищи или иными влияниями. В течение суток поджелудочная железа выделяет примерно 1/5 от запасов имеющегося в ней инсулина.
Главным стимулятором секреции инсулина является повышение концентрации глюкозы в крови выше 5,5 ммоль/л, максимума секреция достигает при 17-28 ммоль/л. Особенностью этой стимуляции является двухфазное усиление секреции инсулина:
- первая фаза длится 5-10 минут и концентрация гормона может 10-кратно возрастать, после чего его количество понижается,
- вторая фаза начинается примерно через 15 минут от начала гипергликемии и продолжается на протяжении всего ее периода, приводя к увеличению уровня гормона в 15-25 раз.
Чем дольше в крови сохраняется высокая концентрация глюкозы, тем большее число β-клеток подключается к секреции инсулина.
Индукция синтеза инсулина происходит от момента проникновения глюкозы в клетку до трансляции инсулиновой мРНК. Она регулируется повышением транскрипции гена инсулина, повышением стабильности инсулиновой мРНК и увеличением трансляции инсулиновой мРНК.
Активация секреции инсулина
1. После проникновения глюкозы в β-клетки (через ГлюТ-1 и ГлюТ-2) она фосфорилируется гексокиназой IV (глюкокиназа, обладает низким сродством к глюкозе),
2. Далее глюкоза аэробно окисляется, при этом скорость окисления глюкозы линейно зависит от ее количества,
3. В результате нарабатывается АТФ, количество которого также прямо зависит от концентрации глюкозы в крови,
4. Накопление АТФ стимулирует закрытие ионных K + -каналов, что приводит к деполяризации мембраны,
5. Деполяризация мембраны приводит к открытию потенциал-зависимых Ca 2+ -каналов и притоку ионов Ca 2+ в клетку,
6. Поступающие ионы Ca 2+ активируют фосфолипазу C и запускают кальций-фосфолипидный механизм проведения сигнала с образованием ДАГ и инозитол-трифосфата (ИФ3),
7. Появление ИФ3 в цитозоле открывает Ca 2+ -каналы в эндоплазматической сети, что ускоряет накопление ионов Ca 2+ в цитозоле,
8. Резкое увеличение концентрации в клетке ионов Ca 2+ приводит к перемещению секреторных гранул к плазматической мембране, их слиянию с ней и экзоцитозу кристаллов зрелого инсулина наружу,
9. Далее происходит распад кристаллов, отделение ионов Zn 2+ и выход молекул активного инсулина в кровоток.
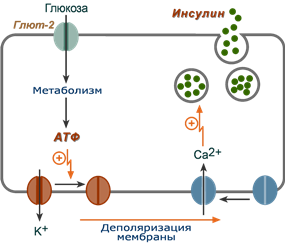
Схема внутриклеточной регуляции секреции инсулина при участии глюкозы
Описанный ведущий механизм может корректироваться в ту или иную сторону под действием ряда других факторов, таких как аминокислоты, жирные кислоты, гормоны ЖКТ и другие гормоны, нервная регуляция .
Из аминокислот на секрецию гормона наиболее значительно влияют лизин и аргинин. Но сами по себе они почти не стимулируют секрецию, их эффект зависит от наличия гипергликемии, т.е. аминокислоты только потенциируют действие глюкозы.
Свободные жирные кислоты также являются факторами, стимулирующими секрецию инсулина, но тоже только в присутствии глюкозы.
Логичной является положительная чувствительность секреции инсулина к действию гормонов желудочно-кишечного тракта – инкретинов (энтероглюкагона и глюкозозависимого инсулинотропного полипептида), холецистокинина, секретина, гастрина, желудочного ингибирующего полипептида.
Клинически важным и в какой-то мере опасным является усиление секреции инсулина при длительном воздействии соматотропного гормона, АКТГ и глюкокортикоидов, эстрогенов, прогестинов. При этом возрастает риск истощения β-клеток, уменьшение синтеза инсулина и возникновение инсулинзависимого сахарного диабета. Такое может наблюдаться при использовании указанных гормонов в терапии или при патологиях, связанных с их гиперфункцией.
Нервная регуляция β-клеток поджелудочной железы включает адренергическую и холинергическую регуляцию. Любые стрессы (эмоциональные и/или физические нагрузки, гипоксия, переохлаждение, травмы, ожоги) повышают активность симпатической нервной системы и подавляют секрецию инсулина за счет активации α2-адренорецепторов. С другой стороны, стимуляция β2-адренорецепторов приводит к усилению секреции.
Также выделение инсулина повышается n.vagus, в свою очередь находящегося под контролем гипоталамуса, чувствительного к концентрации глюкозы крови.
Мишени
Рецепторы инсулина находятся практически на всех клетках организма, кроме нервных, но в разном количестве. Нервные клетки не имеют рецепторов к инсулину, т.к. последний просто не проникает через гематоэнцефалический барьер.
Механизм действия
После связывания инсулина с рецептором активируется ферментативный домен рецептора. Так как он обладает тирозинкиназной активностью, то фосфорилирует внутриклеточные белки — субстраты инсулинового рецептора. Дальнейшее развитие событий обусловлено двумя направлениями: MAP-киназный путь и ФИ-3-киназный механизмы действия (подробно).
При активации фосфатидилинозитол-3-киназного механизма результатом являются быстрые эффекты – активация ГлюТ-4 и поступление глюкозы в клетку, изменение активности «метаболических» ферментов – ТАГ-липазы, гликогенсинтазы, гликогенфосфорилазы, киназы гликогенфосфорилазы, ацетил-SКоА-карбоксилазы и других.
При реализации MAP-киназного механизма (англ. MAP — mitogen-activated protein) регулируются медленные эффекты – пролиферация и дифференцировка клеток, процессы апоптоза и антиапоптоза.
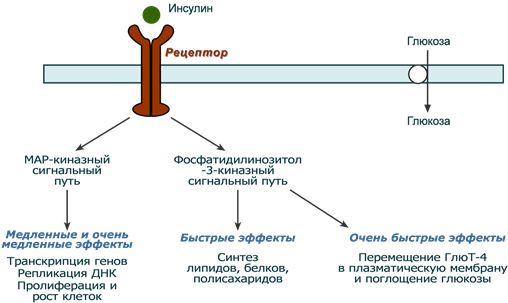
Два механизма действия инсулина
Скорость эффектов действия инсулина
Биологические эффекты инсулина подразделяются по скорости развития:
Очень быстрые эффекты (секунды)
Эти эффекты связаны с изменением трансмембранных транспортов :
1. Активации Na + /K + -АТФазы , что вызывает выход ионов Na + и вход в клетку ионов K + , что ведет к гиперполяризации мембран чувствительных к инсулину клеток (кроме гепатоцитов).
2. Активация Na + /H + -обменника на цитоплазматической мембране многих клеток и выход из клетки ионов H + в обмен на ионы Na + . Такое влияние имеет значение в патогенезе артериальной гипертензии при сахарном диабете 2 типа.
3. Угнетение мембранной Ca 2+ -АТФазы приводит к задержке ионов Ca 2+ в цитозоле клетки.
4. Выход на мембрану миоцитов и адипоцитов переносчиков глюкозы ГлюТ-4 и увеличение в 20-50 раз объема транспорта глюкозы в клетку.
Быстрые эффекты (минуты)
Быстрые эффекты заключаются в изменении скоростей фосфорилирования и дефосфорилирования метаболических ферментов и регуляторных белков.
Печень
- торможение эффектов адреналина и глюкагона (фосфодиэстераза),
- ускорение гликогеногенеза (гликогенсинтаза),
- активация гликолиза (фосфофруктокиназа, пируваткиназа),
- превращение пирувата в ацетил-SКоА (ПВК-дегидрогеназа),
- усиление синтеза жирных кислот (ацетил-SКоА-карбоксилаза),
- формирование ЛПОНП,
- повышение синтеза холестерина (ГМГ-SКоА-редуктаза),
Мышцы
- торможение эффектов адреналина (фосфодиэстераза),
- стимулирует транспорт глюкозы в клетки (активация ГлюТ-4),
- стимуляция гликогеногенеза (гликогенсинтаза),
- активация гликолиза (фосфофруктокиназа, пируваткиназа),
- превращение пирувата в ацетил-SКоА (ПВК-дегидрогеназа),
- усиливает транспорт нейтральных аминокислот в мышцы,
- стимулирует трансляцию (рибосомальный синтез белков).
Жировая ткань
- стимулирует транспорт глюкозы в клетки (активация Глют-4),
- активирует запасание жирных кислот в тканях (липопротеинлипаза),
- активация гликолиза (фосфофруктокиназа, пируваткиназа),
- усиление синтеза жирных кислот (активация ацетил-SКоА-карбоксилазы),
- создание возможности для запасания ТАГ (инактивация гормон-чувствительной-липазы).
Медленные эффекты (минуты-часы)
Медленные эффекты заключаются в изменении скорости транскрипции генов белков, отвечающих за обмен веществ, за рост и деление клеток, например:
1. Индукция синтеза ферментов в печени
- глюкокиназы и пируваткиназы ( гликолиз ),
- АТФ-цитрат-лиазы, ацетил-SКоА-карбоксилазы, синтазы жирных кислот, цитозольной малатдегидрогеназы ( синтез жирных кислот ),
- глюкозо-6-фосфатдегидрогеназы ( пентозофосфатный путь ),
2. Индукция в адипоцитах синтеза глицеральдегидфосфат-дегидрогеназы и синтазы жирных кислот.
3. Репрессия синтеза мРНК, например, для ФЕП-карбоксикиназы ( глюконеогенез ).
4. Обеспечивает процессы трансляции, повышая фосфорилирование по серину рибосомального белка S6.
Очень медленные эффекты (часы-сутки)
Очень медленные эффекты реализуют митогенез и размножение клеток. Например, к этим эффектам относится
1. Повышение в печени синтеза соматомедина, зависимого от гормона роста.
2. Увеличение роста и пролиферации клеток в синергизме с соматомединами.
3. Переход клетки из G1-фазы в S-фазу клеточного цикла.
Инактивация инсулина
Удаление инсулина из циркуляции происходит после его связывания с рецептором и последующей интернализации (эндоцитоза) гормон-рецепторного комплекса, в основном в печени и мышцах. После поглощения комплекс разрушается и белковые молекулы лизируются до свободных аминокислот. В печени захватывается и разрушается до 50% инсулина при первом прохождении крови, оттекающей от поджелудочной железы. В почках инсулин фильтруется в первичную мочу и, после реабсорбции в проксимальных канальцах, разрушается.
Патология
Гипофункция
Инсулинзависимый и инсулиннезависимый сахарный диабет. Для диагностики этих патологий в клинике активно используют нагрузочные пробы и определение концентрации инсулина и С-пептида.
Источник статьи: http://biokhimija.ru/gormony/insulin.html
ИНСУЛИН
Инсулин (лат. insula остров, островок) — гормон поджелудочной железы; относится к группе белковопептидных гормонов.
В 1900 г. Л. В. Соболев доказал, что островки Лангерганса поджелудочной железы (см.) являются местом образования вещества, регулирующего углеводный обмен в организме. В 1921 г. Ф. Бантинг и Бест (С. Н. Best) получили экстракт из островковой ткани поджелудочной железы, содержащий инсулин. В 1925 г. Инсулин был получен в кристаллическом виде. В 1955 г. Сенгер (F. Sanger) изучил аминокислотную последовательность и установил структуру Инсулина крупного рогатого скота и свиней.
Относительная молекулярная масса мономера Инсулин — ок. 6000. Молекула Инсулина содержит 51 аминокислоту и состоит из двух цепей; цепь с N-концевым глицином называется А-цепью и состоит из 21 аминокислоты, вторая — B-цепь — состоит из 30 аминокислот. А- и B-цепи соединены дисульфидной связью, целостность к-рой играет большую роль в сохранении биол, активности молекулы И. (формулу см. ниже).

Наиболее близок по аминокислотному составу к И. человека И. свиньи, молекула к-рого отличается всего лишь на одну аминокислоту в B-цепи (вместо треонина в 30-м положении находится аланин).
Содержание
Биосинтез инсулина, регуляция секреции инсулина
Инсулин синтезируется в базофильных инсулоцитах (бета-клетках) островков Лангерганса поджелудочной железы из своего предшественника — проинсулина. Впервые проинсулин был обнаружен Стайнером (D. F. Steiner) в конце 60-х гг. Проинсулин — одноцепочечный полипептид с относительной мол. массой ок. 10 000, содержит более 80 аминокислот. Проинсулин представляет собой молекулу П., как бы замкнутую пептидом, который был назван соединяющим, или C-пептидом; этот пептид делает молекулу И. биологически неактивной. По иммунологической характеристике проинсулин близок к И. Проинсулин синтезируется на рибосомах инсулоцитов, затем по цистернам цитоплазматической сети молекула проинсулина передвигается к пластинчатому комплексу (комплекс Гольджи), от к-рого отделяются вновь образованные секреторные гранулы, содержащие проинсулин. В секреторных гранулах под действием ферментов от проинсулина отделяется С-пептид и образуется И. Процесс ферментативного превращения проинсулина протекает в. несколько стадий, в результате которых образуется инсулин, промежуточные формы про-инсулина и С-пептид. Все эти вещества обладают разной биол, и иммунной активностью и могут участвовать в регуляции различных видов обмена веществ. Нарушение процессов превращения проинсулина в И. приводит к изменению соотношения этих веществ, появлению анормальных форм И. ив результате этого — к сдвигу в регуляции обмена веществ.
Поступление гормонов в кровь регулируется несколькими механизмами, одним из которых для И. (пусковым сигналом) является повышение содержания глюкозы в крови (см. Гипергликемия); важная роль в регуляции поступления И. принадлежит микроэлементам, гормонам жел.-киш. тракта (в основном секретину), аминокислотам, а также ц. н. с. (см. Гормоны).
Превращение инсулина в организме
При выходе в русло крови часть И. образует комплексы с белками плазмы крови — так наз. связанный инсулин, другая часть остается в форме свободного инсулина. Л. К. Старосельцева и сотр. (1972) установили, что существуют две формы связанного И.: одна форма — комплекс И. с трансферрином, другая — комплекс И. с одним из компонентов альфа-глобулинов сыворотки крови. Свободный и связанный И. отличны друг от друга по биол., иммунным и физ.-хим. свойствам, а также по влиянию на жировую и мышечную ткани, которые являются органами-мишенями и называются инсулинчувствительным и тканями. Свободный И. реагирует с антителами к кристаллическому П., стимулирует поглощение глюкозы мышечной и в какой-то степени жировой тканью. Связанный И. не реагирует с антителами к кристаллическому П., стимулирует поглощение глюкозы жировой тканью и практически не влияет на этот процесс в мышечной ткани. Связанный И. отличается от свободного скоростью метаболизма, поведением в электрофоретическом поле, при гельфильтрации и диализе.
При экстракции сыворотки крови солянокислым этанолом было получено вещество, по биол, эффектам подобное И. Однако это вещество не реагировало с антителами, полученными к кристаллическому Инсулину, и поэтому было названо «неподавляемая инсулиноподобная активность плазмы», или «инсулиноподобное вещество». Изучению инсулиноподобной активности придается большое значение; «неподавляемая инсулиноподобная активность плазмы» многими авторами рассматривается как одна из форм И. Благодаря процессам связывания И. с белками сыворотки крови обеспечивается его доставка к тканям. Кроме того, связанный И. является как бы формой хранения гормона в крови и создает резерв активного И. в русле крови. Определенное соотношение свободного и связанного И. обеспечивает нормальную жизнедеятельность организма.
Количество И., циркулирующего в русле крови, определяется не только скоростью секреции, но и скоростью его метаболизма в периферических тканях и органах. Наиболее активно процессы метаболизма И. протекают в печени. Существует несколько предположений о механизме этих процессов в печени; установлено, что имеются два этапа — восстановление дисульфидных мостиков в молекуле инсулина и протеолиз с образованием биологически неактивных пептидных фрагментов и аминокислот. Существует несколько инсулининактивирующих и инсулиндеградирующих ферментных систем, участвующих в метаболизме И. К ним относятся инсулининактивирующая ферментная система [протеиндисульфидная редуктаза (глютатион)] и инсулиндеградирующая ферментная система, к-рая представлена тремя типами протеолитических ферментов. В результате действия протеиндисульфидной редуктазы происходит восстановление — S— S-мостиков и образование А- и B-цепей И. с последующим протеолизом их до отдельных пептидов и аминокислот. Помимо печени, метаболизм И. происходит в мышечной и жировой тканях, почках, плаценте. Скорость процессов метаболизма может служить контролем за уровнем активного И. и играет большую роль в патогенезе сахарного диабета. Период биол, полураспада И. человека — ок. 30 мин.
Биологическое действие инсулина
Инсулин является универсальным анаболическим гормоном. Один из наиболее ярких эффектов И. — его гипогликемическое действие. И. оказывает влияние на все виды обмена веществ: стимулирует транспорт веществ через клеточные мембраны, способствует утилизации глюкозы и образованию гликогена, ингибирует глюконеогенез (см. Гликолиз), тормозит липолиз и активирует липогенез (см. Жировой обмен), повышает интенсивность синтеза белка. И., обеспечивая нормальное окисление глюкозы в цикле Кребса (легкие, мышцы, почки, печень), способствует образованию макроэргических соединений (в частности, АТФ) и поддержанию энергетического баланса клеток. И, необходим для роста и развития организма (действует в синергизме с соматотропный гормоном гипофиза).
Все биол, эффекты И. самостоятельны и независимы друг от друга, однако в физиологических условиях конечный эффект И. складывается из непосредственной стимуляции биосинтетических процессов и одновременного снабжения клеток «строительным» материалом (напр., аминокислотами) и энергией (глюкозой). Многообразные эффекты И. реализуются путем взаимодействия его с рецепторами клеточных мембран и передачи сигнала (информации) в клетку к соответствующим ферментным системам.
Физиологическим антагонистом Инсулина в регуляции углеводного обмена и обеспечении оптимального для жизнедеятельности организма уровня глюкозы в крови является глюкагон (см.), а также некоторые другие гормоны (щитовидной железы, надпочечников, соматотропный гормон).
Нарушения в синтезе и секреции инсулина могут быть разного характера и иметь различное происхождение. Так, недостаточность секреции И. приводит к гипергликемии и развитию сахарного диабета (см. Диабет сахарный, этиология и патогенез). Избыточное образование И. наблюдается, напр., при гормонально-активной опухоли, исходящей из бета-клеток панкреатических островков (см. Инсулома), и выражается клинически симптомами гиперинсулинизма (см.).
Методы определения инсулина
Методы определения инсулина условно можно разделить на биологические и радиоиммунные. Биол, методы основаны на стимуляции поглощения глюкозы инсулинчувствительными тканями под действием И. Для биол, метода используется диафрагмальная мышца и эпидидимальная жировая ткань, получаемая от крыс чистых линий. Кристаллический И. или исследуемая сыворотка крови человека и препараты диафрагмальной мышцы или эпидидималыюй жировой ткани (лучше изолированные жировые клетки, полученные из эпидидимальной жировой ткани) в буферном р-ре, содержащем определенную концентрацию глюкозы, помещаются в инкубатор. По степени поглощения глюкозы тканью и соответственно убыли ее из инкубируемой среды рассчитывают содержание И. в крови, используя при этом стандартную кривую.
Свободная форма И. усиливает поглощение глюкозы в основном на диафрагмальной мышце, с к-рой практически не реагирует связанная форма И., поэтому, используя диафрагмальный метод, можно определить количество свободного И. Поглощение глюкозы эпидидимальной жировой тканью стимулируется в основном связанной формой И.; но с жировой тканью частично может реагировать и свободный И., поэтому данные, полученные при инкубации с жировой тканью, можно называть общей инсулинной активностью. Физиол, уровни свободного и связанного И. колеблются в очень широких пределах, что, видимо, связано с индивидуальным типом гормональной регуляции обменных процессов, и могут в среднем составлять в норме 150—200 мкед/мл свободного И. и 250—400 мкед/мл связанного И.
Радиоиммунный метод определения И. основан на конкуренции меченого и немеченого И. в реакции с антителом к И. в анализируемой пробе. Количество радиоактивного И., связанного с антителами, будет обратно пропорционально концентрации И. в анализируемой пробе. Наиболее удачным вариантом радиоиммунного метода оказался метод двойных антител, который условно (схематически) можно представить следующим образом. Антитела против И. получают на морских свинках (так наз. антитела первого порядка) и соединяют их с меченым И. (1251). Полученный комплекс повторно соединяют с антителами второго порядка (полученными от кролика). Это обеспечивает стабильность комплекса и возможность реакции замещения меченого И. на немеченый. В результате этой реакции немеченый И. связывается с антителами, а меченый И. переходит в свободный р-р.
Многочисленные модификации этого метода основаны на этапе отделения меченого И. от комплекса с немеченым И. Метод двойных антител положен в основу приготовления готовых наборов для радиоиммунного метода определения И. (фирмами Англии и Франции).
Препараты инсулина
Для медицинских целей Инсулин получают из поджелудочной железы крупного рогатого скота, свиней и китов. Активность И. определяют биол, путем (по способности понижать содержание сахара в крови у здоровых кроликов). За единицу действия (ЕД), или интернациональную единицу (ИЕ), принимают активность 0,04082 мг кристаллического инсулина (стандарта). И. легко соединяется с двухвалентными металлами, особенно с цинком, кобальтом, кадмием, и может образовывать комплексы с полипептидами, в частности с протамином. Это свойство было использовано при создании препаратов И. пролонгированного действия.
По длительности действия различают три типа препаратов И. Препаратом короткого действия (ок. 6 час.) является инсулин отечественного производства (И. крупного рогатого скота и свиней). Препарат средней продолжительности действия (10—12 час.) — суспензия цинк-инсулина аморфного — отечественный препарат, аналогичный зарубежному препарату семиленте. К препаратам длительного действия относятся протамин-цинк-инсулин для инъекций (16—20 час. действия), суспензия инсулин-протамина (18— 24 час.), суспензия цинк-инсулина (до 24 час.), суспензия цинк-инсулина кристаллического (до 30—36 час. действия).
Фармакологическую характеристику наиболее употребляемых препаратов И. и формы их выпуска — см. Гормональные препараты, таблица.
Показания и противопоказания
И. является специфическим противодиабетическим средством и применяется в основном при сахарном диабете; абсолютным показанием является наличие кетоацидоза и диабетической комы. Выбор препарата и его дозировка зависят от формы и тяжести течения болезни, возраста и общего состояния больного. Подбор доз и лечение И. проводится под контролем содержания сахара в крови и в моче и наблюдением за состоянием больного. Передозировка И. грозит резким падением содержания сахара в крови, гипогликемической комой. Конкретные показания к применению тех или иных препаратов И. при сахарном диабете у взрослых и детей — см. Диабет сахарный, лечение.
Препараты И. применяются для лечения некоторых психических болезней. В СССР инсулиношоковое лечение шизофрении было применено в 1936 г. А. С. Кронфельдом и Э. Я. Штернбергом. С появлением нейролептиков лечение И. стало методом выбора — см. Шизофрения.
В небольших дозах И. иногда назначают при общем истощении, фурункулезе, рвоте беременных, гепатитах и др.
Все препараты И. пролонгированного действия вводят только под кожу (или внутримышечно). Внутривенно (напр., при диабетической коме) можно вводить только р-р инсулина кристаллического для инъекций. Нельзя вводить суспензии цинк-инсулина (и другие препараты И. пролонгированного действия) в одном шприце с р-ром инсулина для инъекций; в случае необходимости вводят р-р инсулина для инъекций отдельным шприцем.
Противопоказание — аллергия к Инсулину; относительные противопоказания — заболевания, протекающие с гипогликемией. Необходима осторожность при лечении И. больных, у которых наблюдаются коронарная недостаточность и нарушения мозгового кровообращения.
Библиография: Биохимия гормонов и гормональной регуляции, под ред. Н. А. Юдаева, с. 93, М., 1976; Ньюсхолм Э.И Старт К. Регуляция метаболизма, пер. с англ., с. 387 и др., М., 1977; Проблемы медицинской энзимологии, под ред. G. Р. Мардашева, с. 40, М., 1970, библиогр.; Руководство по клинической эндокринологии, под ред. В. Г. Баранова, Л., 1977; Сахарный диабет, под ред. В. Р. Клячко, с. 130, М., 1974; Старосельцева Л. К. Различные формы инсулина в организме и их биологическое значение, в кн.: Совр. вопр, эндокрин., под ред. H. А. Юдаева, в. 4, с. 123, М., 1972; Юдаев Н. А. Биохимия гормональной регуляции обмена веществ, Вестн. АН СССР, № 11, с. 29, 1974; Banting F. G., а. Вest С. H. Internal secretion of pancreas, J. Lab. clin. Med., v. 7, p. 251, 1922; Cerasi E. a. Luft R. Diabetes mellitus — a disorder of cellular information transmission, Horm. metaboi. Res., v. 4, p. 246, 1970, bibliogr.; Insulin, ed. by R. Luft, Gentofte, 1976; Steiner D. F. a, o. Proinsulin and the biosynthesis of insulin, Recent Progr. Hormone Res., v. 25, p. 207, 1969, bibliogr.
В. С. Ильин, Л. К. Старосельцева
Источник статьи: http://xn--90aw5c.xn--c1avg/index.php/%D0%98%D0%9D%D0%A1%D0%A3%D0%9B%D0%98%D0%9D
Инсулин – популярнейшая
молекула XX столетия
В истории химии случались события, по своему драматизму напоминавшие штурм неприступной вершины, на которую пытаются взойти одновременно независимые группы альпинистов по различным маршрутам. Все это сопровождается обстановкой состязания – кто взойдет на вершину первым?
Далее речь пойдет о синтезе инсулина – событии, ставшем заметным достижением в химической науке. Точно так же, как перед штурмом вершины альпинисты создают базовые, промежуточные и штурмовые лагеря, синтез инсулина был хорошо подготовлен, но не теми, кто вышел на покорение вершины, а основательными работами исследователей-предшественников. Можно уверенно сказать, что создание исходного плацдарма впечатляет не меньше, чем последующий штурм. Инсулин по праву можно назвать популярнейшей молекулой ХХ столетия; с исследованиями этого соединения связаны имена семи (!) нобелевских лауреатов.
Белок, спасающий жизнь
В середине XX в. инсулин был одним из наиболее интенсивно изучаемых веществ. Причина в том, что удалось объяснить происхождение одного из тяжелейших заболеваний – сахарного диабета. Болезнь возникает, когда в организме недостаточно гормона* инсулина. Инсулин запускает процессы, обеспечивающие поступление глюкозы (сахара) в клетки, а также стимулирует внутриклеточные механизмы, позволяющие усваивать глюкозу.
При недостатке инсулина глюкоза не расходуется клетками, она накапливается в крови и начинает через почки поступать в мочу. Повышенный уровень глюкозы в крови и ее выведение с мочой приводят к похуданию, чрезмерному мочеотделению, постоянному ощущению сильной жажды и голода. Организм старается компенсировать дефицит калорий, которые он теряет с мочой в виде глюкозы, и начинает использовать жировые запасы и тканевые белки (главным образом мышечные). Возникают утомление, сонливость, тошнота, нарушаются обменные процессы, что может привести к диабетической коме, а при отсутствии лечения к смерти.
Сахарный диабет встречается среди населения всех стран и у представителей всех рас. Самое раннее описание этого заболевания было сделано примерно 3000 лет назад в Древней Индии. Подробные симптомы болезни (обильное мочеотделение, чрезмерная жажда и потеря веса) были описаны в I в. н.э. Болезнь получила свое название от греческого diabetes, что означает «протекаю, прохожу сквозь» (имеется в виду чрезмерное мочеотделение).
Планомерное изучение этого заболевания длилось не одно столетие. В XVII в. английский врач Т.Уиллис обратил внимание на то, что моча у пациентов с такими симптомами имеет сладковатый вкус (провести подобный анализ мог только истинный ученый). Картина начала проясняться после опытов французского физиолога Клода Бернара (1813–1878), в которых он наблюдал собак с удаленной поджелудочной железой. Его опыты продолжили в 1889 г. немецкие физиологи Йозеф фон Меринг и Оскар Минковский. Они удаляли хирургическим путем поджелудочную железу у собак и затем наблюдали у них резкий подъем концентрации глюкозы в крови, ее появление в моче и другие признаки сахарного диабета. Таким образом они экспериментально доказали связь между поджелудочной железой и сахарным диабетом.
Д.Маклеод
(1876–1935)
Некоторые физиологи высказывали предположение, что поджелудочная железа вырабатывает вещество, которое способствует усваиванию в организме глюкозы. В 1916 г. немецкий физиолог Шарпи-Шафер назвал это гипотетическое вещество инсулин (от латинского insula – островок, поскольку отчетливо наблюдаемые группы клеток поджелудочной железы к этому моменту именовали островками Лангерганса). Тогда это было только предположение, которое впоследствии полностью подтвердилось.
Ф.Бантинг
(1891–1941)
В 1921 г. трое канадских исследователей – профессор физиологии университета в г. Торонто (Канада) Джон Маклеод, врач-хирург Фредерик Бантинг и врач-физиолог Чарлз Бест сумели выделить инсулин из поджелудочной железы подопытных животных. Первые же опыты по введению полученного препарата собакам с удаленной поджелудочной железой продемонстрировали значительное снижение уровня сахара в крови животных и улучшение клинической картины.
Ч.Бест
(1899–1978)
11 января 1922 г. (знаменательный факт в истории мировой медицины) более чистый и активный препарат инсулина был введен первому пациенту – подростку, страдавшему тяжелой формой диабета. После полученного положительного эффекта были проведены аналогичные испытания еще на нескольких пациентах. Возникло новое направление в медицинской науке – гормонотерапия.
В 1923 г. Маклеод и Бантинг были удостоены Нобелевской премии по физиологии и медицине «за открытие инсулина». Бест не был включен в список лауреатов, и Бантинг отдал ему половину полученных денег (жест, достойный истинного ученого).
В 1926 г. было налажено серийное производство инсулина. Многие тысячи больных сахарным диабетом, ранее обреченных на смерть, были спасены и могли вести сравнительно нормальную жизнь, регулярно принимая лекарство.
Физиологи Маклеод и Бантинг использовали для лечения больных экстракт поджелудочной железы животных. Однако химиков всегда интересовало, как именно устроено то или иное соединение. Инсулин в кристаллическом виде впервые сумел получить в 1926 г. Дж.Абель. Именно благодаря его работам удалось наладить промышленное производство препарата. Абель также определил состав инсулина, стало понятно, что вещество представляет собой белковую молекулу. C этого момента исследования инсулина из медицины переходят в область химии, точнее, в руки биохимиков.
Ф.Сенгер
(р. 1918)
Все упомянутые выше работы подготовили решающий этап, позволивший выяснить, как устроена молекула, привлекавшая внимание столь большого числа исследователей. Решить эту задачу удалось американскому биохимику Фредерику Сенгеру. Вначале он разработал способ идентификации концевых аминогрупп в белковой молекуле путем обработки в щелочной среде динитрофторбензолом (впоследствии этот метод стал классическим). Далее он буквально разобрал на части всю молекулу инсулина и определил состав полученных аминокислот с помощью самых современных методов – электрофореза, разработанного А.Тизелиусом (Нобелевская премия, 1948 г.) и хроматографии, усовершенствованной А.Мартином и Р.Сингом (нобелевские лауреаты, 1952 г.). Однако установить, из каких аминокислот собрана белковая молекула, лишь половина дела, притом менее сложная. Главное – выяснить их последовательность в цепи.
Сенгер разработал план, по которому с помощью специально подобранных ферментов (биологических катализаторов) проводил расщепление белковой цепи на небольшие отрезки в заранее намеченных участках, а потом сопоставлял их состав. Работа представляла собой безупречное сочетание логики и экспериментального мастерства, и в 1958 г. ученому была присуждена Нобелевская премия «за работы по структуре протеинов, особенно инсулина». Свой метод Сенгер довел буквально до совершенства, со временем его методика стала общим принципом исследования структуры белков.
Винсент
Дю Виньо
(1901–1978)
Попутно отметим, что Сенгер, применив похожие логические построения, но несколько изменив методику и используемые реагенты, сумел установить последовательность фрагментов в структуре знаменитой двойной спирали ДНК. За эти исследования в 1980 г. Сенгеру (совместно с У.Гилбертом и П.Бергом) была присуждена еще одна Нобелевская премия «за вклад в определение последовательности оснований в нуклеиновых кислотах». Таким образом, Сенгер – единственный дважды нобелевский лауреат по химии. Никто не мог предположить, что эти исследования ДНК со временем откроют новую страницу в химии инсулина, но об этом речь пойдет несколько позже.
Дороти
Кроуфут-Ходжкин
(1910–1994)
Американский биохимик Винсент Дю Виньо, в течение нескольких лет изучавший инсулин, узнав о работах Сенгера, решил воспользоваться его методикой для расшифровки структуры двух других гормонов (вазопрессина и окситоцина). Однако он не только установил строение, но и синтезировал молекулы этих гормонов. Фактически он был первым, кто сумел синтезировать природные полипептиды. Эта работа ученого была отмечена Нобелевской премией в 1955 г., т.е. он получил премию на три года раньше Сенгера, чьи идеи помогли ему добиться столь великолепного результата. Работы Дю Виньо фактически открыли дорогу к синтезу инсулина.
Тем временем изучение инсулина продолжалось. Исследование лечебных свойств инсулина позволило установить, что его цинковый комплекс из нескольких молекул, так называемый Zn-инсулин, обладает более длительным лечебным действием. Строение этого комплекса оказалось весьма сложным (он содержит почти 800 атомов), поэтому были привлечены физико-химические методы анализа. В 1972 г. английский биофизик Дороти Кроуфут-Ходжкин (лауреат Нобелевской премии 1964 г. за определение с помощью рентгеновских лучей структур биологически активных веществ) установила трехмерную структуру этого необычайно сложного комплекса.
Упрощенный язык биохимиков
Прежде чем рассмотреть строение молекулы инсулина, познакомимся с тем, как биохимики изображают молекулы белков.
Все белки представляют собой полимеры, цепи которых собраны из фрагментов аминокислот. Аминокислоты – это органические соединения, содержащие в своем составе аминогруппу NH2 и карбоксильную группу СООН. В образовании белков участвуют только такие аминокислоты, у которых между аминогруппой и карбоксильной группой всего один углеродный атом. В общем виде они могут быть представлены формулой H2N–CH(R)–COOH. Группа R, присоединенная к атому углерода (тому, который находится между амино- и карбоксильной группой), определяет различие между аминокислотами, образующими белки. Эта группа может состоять только из атомов углерода и водорода, но чаще содержит помимо С и Н различные функциональные группы. Из всего многообразия существующих аминокислот (теоретически количество возможных аминокислот неограниченно) в образовании белков участвуют только двадцать, так называемые «фундаментальные» аминокислоты. Для «строительства» инсулина природа использовала 16 аминокислот (из допустимых двадцати) (табл.1).
Аминокислоты, участвующие в создании инсулина
| Название | Структура | Обозначение* |
| Глицин |  | Гли |
| Аланин |  | Ала |
| Валин |  | Вал |
| Лейцин |  | Лей |
| Изолейцин |  | Иле |
| Серин |  | Сер |
| Цистеин |  | Цис |
| Лизин |  | Лиз |
| Аргинин |  | Арг |
| Аспарагин |  | Асн |
| Глутаминовая кислота |  | Глу |
| Глутамин |  | Глн |
| Фениаланин |  | Фен |
| Тирозин |  | Тир |
| Гистидин |  | Гис |
| Пролин |  | Про |
* В международной практике принято сокращенное обозначение перечисленных аминокислот с помощью латинских трехбуквенных сокращений, например глицин – Gly, аланин – Ala и т.п.
Белковая молекула образуется в результате последовательного соединения аминокислот, при этом карбоксильная группа одной кислоты взаимодействует с аминогруппой соседней молекулы, в результате образуется пептидная связь –CO–NH– и выделяется молекула воды. На схеме 1
(см. с. 6) показано последовательное соединение аланина, валина и глицина.

Из превращений, показанных на схеме 1, следует, что при любом количестве соединяемых аминокислот на одном конце возникшей цепочки обязательно будет находиться аминогруппа, а на другом – карбоксильная. Фрагменты соединенных аминокислот обозначены (под фигурными скобками) теми сокращенными буквосочетаниями, которые указаны в табл. 1. Таким образом, вместо структурной формулы мы можем использовать сокращенное обозначение получившегося трипептида: ала-вал-гли. Поскольку количество аминокислот, используемых природой, всего двадцать, то подобные сокращения позволяют компактно записать формулу любого белка, и никакой неясности при этом не возникнет.
Молекула инсулина, как установил Сенгер, состоит из 51 аминокислотного остатка (это один из самых короткоцепочечных белков) и представляет собой две соединенные между собой параллельные цепи неодинаковой длины. На схеме 2 показана последовательность аминокислот в молекуле инсулина: А-цепь содержит 21 аминокислотный остаток, Б-цепь – 30.

Содержащиеся в молекуле остатки аминокислоты цистеина (сокращенное обозначение Цис) образуют дисульфидные мостики S-S-, которые связывают две полимерные цепи молекулы и, кроме того, образуют перемычку внутри А-цепи. При таком компактном изображении белковой молекулы символы химических элементов используют только для обозначения дисульфидных мостиков и концевых групп (NH2 и COOH).
Для сравнения далее показана структурная формула инсулина в виде объемной шаростержневой модели (схема 3).

Согласитесь с тем, что биохимики выбрали компактный и необычайно удобный для написания способ изображения белковых молекул.
От демонтажа к сборке
Казалось бы, после того как установлена структура молекулы, синтезировать ее заново не составит большого труда.
Основная трудность при сборке белковой молекулы – добиться, чтобы необходимые аминокислоты соединялись в строго определенном порядке. При этом нужно учитывать, что аминокислота способна реагировать не только с другой аминокислотой, но и сама с собой, и в итоге может получиться молекула, не имеющая ничего общего с тем, что синтезирует живой организм.
К моменту, когда решался вопрос о синтезе инсулина, было разработано несколько соответствующих методик. Для того чтобы аминокислота, которую намечено было присоединить к растущей цепи, не реагировала сама с собой, ее реакционноспособные концы (аминогруппу NH2 и карбоксильную группу СООН) блокировали специальным образом: карбоксильную группу переводили в п-нитрофениловый эфир, а со стороны аминогруппы присоединяли карбоксибензильную группу. Такая блокированная молекула реагировала с аминогруппой, находящейся на конце растущей цепи, по схеме 4 (cм. c. 8).

В результате растущая цепь удлинялась на одно пептидное звено. Однако теперь на конце цепи разместилась блокирующая карбоксибензильная группа. Чтобы сделать «аминный хвост» реакционноспособным, т. е. перевести его в активную форму, осуществляли обработку бромоводородом с уксусной кислотой по схеме 5 (cм. c. 8).

В результате аминогруппа на конце цепи (она показана в виде аммониевой соли с HBr) вновь была готова реагировать с очередной аминокислотой (естественно, тоже содержащей блокирующие группы). Параллельно были разработаны и другие методы сборки полипептидных цепей.
К полному синтезу инсулина в 1962 г. приступили практически одновременно три группы исследователей: группа П.Катсоянниса в г. Питсбурге (США), группа Г.Цана в г. Аахене (Германия), а также группа китайских химиков (Шанхай и Пекин). Все три группы действовали по весьма похожим стратегиям: собрали отдельно короткую и длинную цепи из заготовленных фрагментов, а затем соединяли обе цепи дисульфидными мостиками.
Аахенская группа Г.Цана (он в центре)
Короткую А-цепь все три группы исследователей собирали из двух одинаковых блоков.
1-й блок: гли-иле-вал-глу-глн-цис-цис-тир-сер;
2-й блок: иле-цис-сер-лей-тир-глн-лей-глу-асн-тир-цис-асн.
Длинную Б-цепь собирали из четырех полипептидных блоков, однако длина этих блоков у разных групп ученых несколько различалась (табл. 2).
Полипептидные блоки для сборки Б-цепи инсулина
| Исследова- тельская группа | 1-й блок | 2-й блок | 3-й блок | 4-й блок |
| Аахенская | Фен-вал-асн-глн- гис-лей-цис-гли | Сер-гис-лей- вал-глу-ала | Лей-тир-лей- вал-цис-глу | Глу-арг-глу-фен-фен- тир-тир-про-лиз-тир |
| Питсбургска | Фен-вал-асн-глн- гис-лей-цис-гли-сер | Гис-лей-вал-глу | Ала-лей-тир-лей- вал-цис-глу | Глу-арг-глу-фен-фен- тир-тир-про-лиз-тир |
| Пекинско- шанхайская | Фен-вал-асн-глн- гис-лей-цис-гли | Сер-гис-лей-вал- глу-ала-лей-тир | Лей-вал-цис-глу | Глу-арг-глу-фен-фен- тир-тир-про-лиз-тир |
Различия возникли из-за того, что методы соединения блоков и способы промежуточной защиты, используемые каждой из исследовательских групп, были неодинаковы. Естественно, на последнем этапе у всех групп получились одинаковые цепи. Приблизительно год ушел на создание исходных блоков. Подстегиваемая обстановкой соревнования аахенская группа интенсифицировала работу и в декабре 1963 г. сообщила об успешном синтезе инсулина. Эта группа буквально вырвала первенство у питсбургских химиков, которые сообщили об успешном результате в марте 1964 г. Окончательный выход чистого продукта колебался в пределах 0,02–0,07%. У китайских химиков выход был несколько выше (1,2–2,5%); разумеется, о производстве инсулина по таким методикам не могло быть и речи.
Питсбургская группа П.Катсоянниса
(он второй справа)
Синтез инсулина стал убедительной победой классической синтетической химии пептидов. Несмотря на низкий выход продукта, все признавали, что была проделана выдающаяся работа, которая позволила изменить образ мышления химиков, сформулировать новые принципы сборки больших молекул, отработать стратегию синтеза и подобрать оптимальные методики. Все это заметно повысило общий уровень органической химии. Тем не менее истинного триумфа не получилось, потому что почти одновременно с успешным завершением этих работ появилась принципиально иная, более совершенная методика сборки белковых молекул.
Главное – закрепить хвост
Профессор Рокфеллеровского университета (Нью-Йорк) Роберт Меррифилд, занимаясь химией белков, высказал оригинальную идею: первую аминокислоту можно закрепить одним концом на некой нерастворимой поверхности (носителе). Затем следует присоединить к другому ее концу следующую аминокислоту, при этом нежелательные побочные продукты и промежуточные реагенты, не вступившие в реакцию, можно будет вымывать из реакционного сосуда после каждой стадии, а растущий полипептид, прикрепленный к носителю, останется при этом незатронутым. Молекулы растущих полипептидов будут подвешены «за хвост» к твердой поверхности носителя, а когда процесс синтеза завершится, конечный полипептид можно отделить от носителя.
Меррифилду удалось реализовать эту идею. Первую аминокислоту присоединяют к нерастворимому полимерному гелю (сшитый полистирол) с введенными в него хлорметильными группами CH2Cl, которые способны реагировать с СООН-группами аминокислоты. Чтобы взятая для реакции аминокислота не прореагировала сама с собой и не присоединилась аминогруппой к подложке, NH2-группу этой кислоты предварительно блокируют объемистым заместителем – [(С4Н9)3]3ОС(О)-группой. После того как аминокислота присоединилась к полимерной подложке, блокирующую группу удаляют и в реакционную смесь вводят другую аминокислоту, у которой также предварительно заблокирована NH2-группа. В такой системе возможно только взаимодействие NH2-группы первой аминокислоты и COOH-группы второй аминокислоты, которое проводят в присутствии катализаторов (солей фосфония). Далее схему присоединения повторяют, вводя третью аминокислоту. Вся схема синтеза полипептидных цепей, позволяющая чередовать аминокислотные остатки в заданном порядке, выглядит следующим образом (схема 6).

На последней стадии полученные полипептидные цепи отделяют от полистирольной подложки действием HBr в присутствии трифторуксусной кислоты F3CCOOH.
Р.Меррифилд
(р. 1921 г.)
Меррифилд не только экспериментально проверил эффективность предложенного метода, но и сконструировал аппарат, который практически автоматизировал пептидный синтез. Это устройство представляло собой контейнер для аминокислот и реагентов – реакционный сосуд с автоматическими впускным и выпускным клапанами и программным механизмом, который регулировал последовательность процессов и длительность каждой стадии.
С помощью сконструированного аппарата Меррифилд и его коллеги синтезировали инсулин всего за 20 дней (притом с выходом в десятки процентов), в то время как «первопроходцы» – аахенская, питсбургская и шанхайская группы – затратили на это больше года.
В 1985 г. Меррифилд был удостоен Нобелевской премии «за развитие методологии твердофазного химического синтеза».
Во время проведения описанных выше работ химиков не оставляла мысль, что те задачи, которые ученые решают с таким трудом, Природа решает легко и исключительно аккуратно. Синтез белков в живых организмах проходит в мягких условиях, быстро и без образования побочных продуктов. До определенного момента химики могли лишь с удивлением и интересом наблюдать подобные «синтезы», однако стремительное развитие биохимии позволило активно вмешаться в эти процессы, в том числе открыть принципиально новый способ синтеза инсулина.
Ранее было сказано, что Ф.Сенгер (установивший структуру инсулина) сумел определить последовательность фрагментов в структуре ДНК, за что был удостоен второй Нобелевской премии. Эта работа позволила биохимикам перейти к следующему этапу – встраивать в генетический код ДНК заранее намеченные фрагменты. Основная идея состояла в том, чтобы в ДНК некоторых бактерий включать гены высших организмов. В результате бактерии приобретают способность синтезировать соединения, которые прежде могли синтезировать только высшие организмы. Такая технология получила название «генная инженерия».
В 1981 г. канадский биохимик Майкл Смит был приглашен в научные соучредители новой биотехнологической компании «Зимос». Один из первых контрактов фирмы был заключен с датской фармацевтической компанией «Ново» по разработке технологии производства человеческого инсулина в дрожжевой культуре. В результате совместных усилий инсулин, полученный по новой технологии, в 1982 г. поступил в продажу. В 1993 г. за цикл работ в этой области М.Смит (совместно с К.Муллисом) получил Нобелевскую премию. В настоящее время инсулин, получаемый методом генной инженерии, практически вытеснил инсулин животных.
М.Смит (1932–2000)
Итак, мы познакомились с четырьмя способами получения инсулина: экстракцией из поджелудочной железы животных (группа Д.Маклеода), многоступенчатым синтезом (группа Г.Цана), автоматизированной сборкой (Р.Меррифилд), методом генной инженерии (М.Смит). Оставим в стороне медицинский аспект проблемы, сосредоточим внимание на химии. Могло сложиться впечатление, что работы Смита сделали ненужными все предшествующие исследования. На самом деле это не так, все методы неразрывно связаны, ни один из этапов упомянутых исследований нельзя «выбросить». Инсулин, выделенный из поджелудочной железы животных, позволил Сенгеру определить его структуру, а без этого последующий синтез был невозможен. Группа Цана разработала химические приемы сборки цепей и способы промежуточной блокировки функциональных групп, которыми воспользовался Меррифилд при создании автоматической установки синтеза. Работы Смита, по существу, опирались на весь предшествующий опыт, накопленный при изучении инсулина. При синтезе некоторых короткоцепочечных гормонов автоматическая установка Меррифилда технически оказалась предпочтительнее «генной инженерии».
Обобщая, можно сказать, что все этапы, которые мы рассмотрели, – это естественный, традиционный и, если не бояться торжественных слов, величественный путь науки.
* Гормоны (от греческого  – привожу в действие, побуждаю) – специфические физиологически активные вещества, вырабатываемые специальными эндокринными органами или тканями, секретируемые в кровь или лимфу и действующие на строение и функции организма.
– привожу в действие, побуждаю) – специфические физиологически активные вещества, вырабатываемые специальными эндокринными органами или тканями, секретируемые в кровь или лимфу и действующие на строение и функции организма.
Источник статьи: http://him.1sept.ru/article.php?ID=200800801